Antonio Chávez Sánchez Silva, escritor autodidacta
Correo-e: hnc.correo@gmail.com
(PARTE 1) (a)
Resumen
Entre tanto, para efectos de aclarar algunos principios argumentales en este ensayo:
- Se asume que cualquier entidad sobrenatural de significado y definición religiosa es un agente sobrenatural personificado con el que interactúan de modo específico las personas individual o colectivamente (plegarias, sacrificios, ritos, dirigidos a ellos esperando algo a cambio: Dios -Yahvé, Alá-, dioses~deidades, ángeles, ancestros, espíritus, etc.), y este agente está más o menos involucrado en la observación de la conducta de sus seguidores y/o el control del mundo que les rodea.
- Estas son representaciones agentivas cuya descripción, definición y personalidad están construidas culturalmente, y son culturalmente transmitidas, adquiridas y asimiladas al esquema cognitivo del individuo. Se rechaza pues el concepto «gen de Dios» (Hamer 2004; cf. Zimmer 2004). (Sin embargo en la segunda parte se analizará la supuesta herencia genética de la religiosidad, y en la tercera parte la supuesta causación de la prosocialidad por la religión.)
- Cuando se sugiere que «la religión es natural» (Bloom 2007; en contraste p.ej. con el ateísmo), se hace referencia a que existen predisposiciones cognitivas que ayudan de modo más o menos directo a una, a veces extremadamente, fácil adquisición de los conceptos culturales religiosos (ver también Banerjee & Bloom 2013). No se hace referencia al concepto de que la religión es innata, ni a que exista algún aspecto genético-neural enfocado exclusivamente a ella. Aunque parezca confuso el tratamiento innatista y evolutivo que se le da a la agencia en la religión, debe estar claro que un asunto es la atribución de intención o de diseño y otra la creencia en dioses.

Esquematización gráfica de la propuesta. Durante la percepción, en milésimas de segundo, los conceptos de origen antiguo evolutivo y socialmente relevantes intercambian contenidos para completar la información entrante ambigua o abrumadora. Por ejemplo, varias circunstancias problemáticas presentándose a la vez sin tener causas evidentes, son codificadas en una primera fase computacional cruzando la causalidad intencional y la construcción de posibles escenarios contextuales. El proceso incluye la activación de emociones. Se considera que esto es lo que simplemente se llama pensamiento mágico, de carácter intrínsecamente sobrenatural. En una segunda fase, sin embargo, esta información autogenerada interactúa con la memoria y el aprendizaje cultural, se construye narrativa para formar un juicio (una creencia) y se activan procesos inhibitorios y analíticos, que sin embargo pueden relajarse bajo la influencia de la dopamina. De esto depende que el individuo tome una decisión afirmativa o negativa respecto a una creencia mágica, como explicación ya consciente y verbalizable.
(Hacer click en la imagen para ampliarla)

De los no muchos estudios pertinentes, se ha mostrado que el volumen del hipocampo está menos asociado a regulación genética (Sullivan et al. 2001). Esto sugiere que el entorno puede influir fuertemente en su estructura anatómica, consistentemente con sus capacidades neuroplásticas y su papel central en la consolidación de la memoria y el aprendizaje (Toga & Thompson 2005 p. 10). Esto a su vez es consistente con el papel de esta región temporomedial del cerebro en aspectos como el mental time travel (MTT), puesto que es necesario que sea funcionalmente susceptible de modulación de acuerdo a los cambiantes estímulos externos que deben ser metarrepresentados espaciotemporalmente. P.ej. la maduración del MTT hacia una consciencia del tiempo dividido en períodos fijos implica la asimilación de conceptos culturales como el calendario (cf. Berntsen & Jacobsen 2008; Suddendorf 1994). Análogamente, este rasgo hipocampal tiene una especial importancia en el desarrollo de las creencias religiosas, en tanto que se conecta recíprocamente con el córtex prefrontal dorsolateral, cuya área frontopolar (área 10 de Brodmann) sustenta los mayores grados de abstracción cognitiva (Badre et al. 2009).2 Así pues, «la arquitectura psicológica responsable de la cognición religiosa es mejor vista como un mental time travel autoilusorio» (Bulbulia 2009 aquí o aquí), en tanto que existe una extrema facilidad confabulatoria para racionalizar y auto justificar simples inferencias, posibilidades meramente imaginarias, o por falta de acceso a procesos de hecho inconscientes y emocionales que soportan las elaboraciones lógico-narrativas en la autoconvicción o creencias «lógicas». En efecto, una confluencia de aspectos mnémicos, emocionales y de automonitoreo soportados por el córtex temporomedial, inferoparietal y prefrontal (también el correlato neural del MTT), con predominancia prefrontal izquierda en casos extremos (delirios), es el «intérprete» del cerebro, «un mecanismo para hacer interpretaciones y formar hipótesis» (Miller & Valsangkar-Smyth 2005; ver «EL HEMISFERIO IZQUIERDO: PATRONES, HIPÓTESIS E IDEAS DELIRANTES»).
La región temporomedial juega un papel importante en la consciencia espacial en conjunción con el córtex parietal medial/posterior (mientras la primera provee un sentido alocéntrico del espacio, el segundo provee uno egocéntrico), aportando la elaboración de un auténtico «mapa» donde se «ubican» representaciones de objetos, conceptos y recuerdos, probablemente en función de cómo sus «células de lugar» representan el espacio físico (Bird & Burgess 2008). Adicionalmente, el hipocampo está fuertemente conectado al córtex cingulado, formando ambos parte de un circuito que incluye los cuerpos mamilares y el tálamo, denominado Circuito Papez. Éste fue originalmente bautizado como el «cerebro visceral» en 1911 (Besada 2010 p. 135) y forma parte del llamado «Sistema Límbico», habiéndose extendido posteriormente para incluir la amígdala y el córtex prefrontal. Es aquí de interés que, estudiando la genética neurofisiológica de la depresión (el SLC6A4, gen transportador de serotonina), se ha encontrado que el acoplamiento amígdalo-prefrontomedial depende de la variación de este gen (Heinz et al. 2004). Ciertamente, se trata de un sistema no solo de representación visceral, sino de la emoción. Como analizado previamente respecto al córtex cingulado anterior (CCA), todo este sistema juega un papel importante en la religión, precisamente como soporte emocional (que es el aspecto que enfatiza p.ej. Ramachandran).
Nótese entonces cómo una representación agentiva (p.ej. Dios) puede en efecto poseer una dimensión más o menos emocional, una gradación de imputación emocional, y cómo esto mismo depende de qué tan abundantes sean las conexiones (o la abundancia de neuromoduladores) dentro del Circuito Papez. Aunque de hecho juegan un papel importante la lectura de símbolos religiosos de fuerte significado socioemocional (de lo que hay evidencia sugerente de tener un rol en la religión: Alcorta & Sosis 2005) o la indoctrinación (es decir el ambiente, que juega un evidente papel en tanto ya se comentó la neuroplasticidad hipocámpica y su menor regulación genética), la predisposición a imbuir de mayor o menor profundidad emocional un proceso cognitivo, adquiere una importancia primaria. Una ilustración de esto es que un aspecto específico como la meditación religiosa está soportada por una compleja interacción, neuroquímicamente controlada, entre el hipocampo y el córtex (cf. Newberg & Iversen 2003 p. 286): lo más probable es que tal control sea un rasgo fenotípico (neurofisiológico) variable entre individuos, y adquisiciones culturales como una específica técnica en meditación (que por cierto bien puede tener significado religioso como no tenerlo) simplemente la modulen.
El cuerpo calloso en su región anterior, denominada genu y que interconecta los lóbulos frontales izquierdo/derecho, es más susceptible a las influencias ambientales que p.ej. su región posterior (que interconecta los lóbulos parietales), «quizás reflejando la prolongada maduración del córtex frontal bien entrada la adultez» (Toga & Thompson 2005 p. 11, respecto a estudios de Pfefferbaum et al. 2001; Sowell et al. 1999; Gogtay et al. 2004). Este es un dato que resulta interesante respecto al natural desbalance prefrontal hacia la derecha en cuanto al sistema dopaminérgico y el pensamiento mágico (ver aquí) y hacia la izquierda en cuanto a interpretaciones narrativas como se comentó arriba. Nuevamente, resulta plausible pensar que la interrelación entre tales aspectos neurocognitivos puede ser fácilmente modulada socioculturalmente, p.ej. para así canalizar la funcionalidad prefrontal respecto a la evaluación de creencias hacia un esquema religioso dado.
Precisamente, antes en «EL VIAJE MENTAL EN EL TIEMPO (‘MENTAL TIME TRAVEL’) Y LA COGNICIÓN RELIGIOSA», se refirieron estudios mostrando que el grado de influencia genética varía en diversas regiones del cerebro. Resulta de fundamental interés que haya mayor tal influencia en lo que respecta al sistema espacial y motor (fronto-parietal), visual (occípito-temporal) y límbico (Schmitt et al. 2008 pp. 1742-1744). También, como de esperarse, Toga & Thompson (2005) mostraron que las clásicas áreas del lenguaje (Broca: habla, Wernicke: comprensión) tienen similares características neuro-genéticas, mientras que, por otro lado las regiones sensoriomotora y occípito-parietal, pero no frontal, aparecieron adicionalmente más influenciadas genéticamente en mellizos que en gemelos (ibid. p. 10); lo que indica que dentro de estas regiones mismas hay grados de asociación genética. Del mismo artículo sobre el MTT hay que recordar necesariamente que el conectoma o materia blanca, esto es el conjunto de fibras neurales que viajan de una región a otra y de un hemisferio a otro del cerebro, conformando un muy denso y complicado sustrato de auténticos cables orgánicos, muestra un patrón de incremento de la influencia genética de un menor grado en la parte anterior o frontal del cerebro, hacia un mayor grado en la posterior, justamente en el córtex parietal y visual (occipital) (Chiang et al. 2009), mientras que la interconexión entre el córtex parietal izquierdo y derecho también está bajo mayor influencia genética que el córtex frontal de ambos hemisferios (ver también la Fig. 3 del artículo mencionado).
Al respecto, se pude notar fácilmente que la selección natural ha resultado en un sistema ‘discriminación visual → acción motora’ rápido y efectivamente adaptativo a costa de interesantes errores como la ilusión óptica o los exagerados sesgos agentivos/causales bajo ciertas circunstancias, como ante la sensación de pérdida de control. Efectivamente, p.ej. respecto al grosor cortical se ha encontrado que el córtex visual primario (V1) tiene fuertes correlaciones genéticas con las regiones sensoriales posteriores y las áreas motoras; mientras la región temporal anterior correlaciona con la frontal anterior pero no con V1; y la frontal media con la parietal inferior (Rimol et al. 2010). En la segunda parte de este artículo se argumentará esta perspectiva de fundamentación biológica mediante el análisis de una demostrada «motivación implícita de control», que es lo que, en última instancia, nos predispone inadvertida, cotidiana y espontáneamente hacia el pensamiento mágico (Pronin et al. 2006), independientemente del marco sociocultural, geográfico, de edad, o de la educación científica (cf. Shweder 1977; Diaconis 1986; Harris 1997; Rosengren & Johnson 2000; Elkan 2001; Schultheiss & Brunstein 2010; Passanisi et al. 2017; Kiken 2019).
Se ha notado previamente (aquí y aquí) que el cerebro humano (con claros indicios filogenéticos observados en primates) muestra especializaciones lateralizadas para la conciencia espacial (hemisferio derecho) y el lenguaje (izquierdo), que responden a específicos tractos conectivos perisilvianos. En conjunto esto lo vengo sugiriendo como base neurocognitiva-evolutiva de la agentividad sobrenaturalista en tanto que consiste en un traslapado entre cognición espacial y social: atribución de significado social y emocional al espacio exterior (‘mente inmaterial’). Pues bien, este sistema perisilviano resulta correlacionar con la red por defecto (infra la Fig. 2 nos ilustra la correspondencia). En efecto, se trata de un asunto muy sugerente puesto en relieve también en ese artículo y en otro más reciente, la importancia que tiene para la agencia y el MTT la red por defecto: un circuito temporoparietal-frontomedial que tiene notoria actividad cuando no realizamos ninguna tarea mental o aún mientras estamos en reposo. Esto, sumado al amplio traslapado entre agencia/MTT/red por defecto sugiere que la agentividad es un proceso fundamentalmente automático (involuntario: Dennett 2011), espontáneo y constante.
Ciertamente, va a ser inevitable agentivizar mucho del mundo que nos rodea, cuando no enteramente y con alcances trans-espaciotemporales (debido a ese traslapado), tal como pasa en muchas cosmovisiones religiosas. Una función neurocognitiva de tal naturaleza compulsiva debería estar bajo regulación genética: en efecto, la red por defecto muestra una doblemente compleja influencia genética, tanto para la conectividad entre las subregiones que la componen, como para la variación neuroanatómica de las subregiones mismas (Glahn et al. 2010). Con un considerable muestreo de 333 individuos pertenecientes a árboles genealógicos vinculados (lo que otorga mayor precisión a la identificación del grado de herencia de los rasgos cognitivos), los autores concluyen que «factores genéticos, más que ambientales, influencian la conectividad a través de la red» (ibid. p. 1226). Este estudio abre las puertas al trabajo de identificar los precisos genes causales aquí, como p.ej. sobre el eje tálamo-cortical (Lokmane & Garel 2011). Luego de algunos comentarios retomaré este punto.


Si enfocamos a esas regiones del cerebro social como estando bajo mayor control del genoma puesto que son el asiento de las capacidades locomotrices y comunicativas del hombre, se hace más fácil aún ver lo estratégicamente adaptativo de ello —todo lo que tiene que ver pues con eventos evolutivos cruciales como la bipedestación y el origen del habla. Pero más importante aquí es que, y me parece que subestimado a diferencia del valor dado al caminar erguido y al habla mismo, el circuito fronto-parietal no solo es la base de la cognición espacial, sino que dado que este proceso también se vincula a los procesos de categorización visual del córtex temporal, todo esto determina que veamos el «espacio externo como agente intencional». Hay muy buenas razones para pensar que venimos con un «numenware», un cableado neural que sostiene lo que sería un reflejo de atribución y detección de patrones, causalidad, intención y continencia de mensajes socioemocionales en el espacio que nos rodea. Esto es algo más que un efecto inevitable, un subproducto o una exaptación del carácter hipertrófico de la conducta social humana: parece que tenemos programada neuro-genéticamente esta espacio-agentividad, pero el significado evolutivo-adaptativo de ello aún es discutido. Esto debería implementarse en la teoría exaptacionista de la religión (que ésta es un subproducto evolutivo), en lugar de interpretarse como una teoría adaptacionista de la religión.
Ya que las referencias que usamos en nuestros artículos son en su 99% en inglés, ayudará saber que F. Rubia (catedrático de la Universidad Complutense y quizás el más importante divulgador neurocientífico en nuestro idioma) sugiere que «la espiritualidad parece ser algo inherente al ser humano desde que se pueden provocar experiencias espirituales o místicas estimulando determinadas regiones cerebrales.» Ciertamente, esto es algo bastante consistente con el estudio de Urgesi et al. (2010), quienes lograron aislar el córtex parietal posterior/inferior como el sustrato de la autotranscendencia o espiritualidad.3 Sugiero el concepto de «modularidad» en esto, apoyándome además en la alta similitud del grado de espiritualidad en gemelos y el probable factor neuroquímico (serotonina) asociado, como los autores notan. En otras palabras, la espiritualidad es pues inherente al ser humano y aquí es entendida como parte de la agentividad (el espacio como agente): tendría una base genética en tanto que sabemos de un fuerte grado de influencia genética en el córtex parietal. Siguiendo a Rubia y como se sostiene en nuestros artículos, el asunto de conceptualizar las experiencias espirituales atañe en cambio a aspectos de aprendizaje sociocultural, y se trata por tanto de un asunto de adquisición no biológica: «si es cierto que no existe religión sin espiritualidad, sí existe, por lo contrario, espiritualidad sin religión».
Tengo que hacer un paréntesis respecto a eso de «modularidad». Tomando como ejemplo a Chavez et al. 2010: «los estudios empíricos también han dado cabida a la hipótesis de que las funciones cerebrales dependen de la coordinación de un mosaico de regiones dispersas del cerebro funcionalmente especializadas (módulos), formando una estructura similar a una red de ensambles neuronales [...] Un módulo se define usualmente como un subconjunto de unidades dentro de una red, de manera que las conexiones entre ellos son más densas que las conexiones con el resto de la red.» Entre tanto, dada una clasificación de la conectividad neural en «estructural» (conexiones físicas como sinapsis y el cableado que conforma la materia blanca), «funcional» (patrones de interacción dinámica) y «efectiva» (efectos causales entre elementos neurales), «mientras que la conectividad estructural (al menos en escalas de tiempo de segundos a minutos) se mantiene bastante estable, la conectividad funcional y efectiva pueden someterse a una rápida reconfiguración del orden de cientos de milisegundos en el curso de la actividad espontánea o evocada mediante tareas» (Sporns 2010 p. 9). Esto puede darnos una idea de cómo el sistema de agencia, claramente modular, puede funcionar mecánicamente disparando outputs de categorización perceptual a nivel de conexión puramente estructural (p.ej. con cierta información sensorial del entorno construir/disparar una atribución de mentalidad), y sin embargo este mismo mecanismo no verse paralizado por la construcción de outputs mucho más elaborados o dependientes del aprendizaje, ya de naturaleza conectiva funcional o efectiva dado que implican la interacción entre varios módulos (p.ej. agencia + memoria + emoción + narrativa en el caso de asimilar una nueva creencia).
Con esto pretendo ilustrar, por un lado, que hay una influencia genética sobre la estructura física de la conectividad (de hecho es algo demostrado), pero que puede decrecer según el tipo, nivel de complejidad o la organización jerárquica de la conectividad (funcional y efectiva) y depender más fuertemente de la influencia del medio. Por otro lado, todo esto explica muy bien cómo es que un mismo cerebro podría internamente sostener esquemas de creencias dispares y cómo es que la investigación psicológica ha logrado identificar pensamiento mágico o conductas supersticiosas (‘espectro agentivo-sobrenaturalista’) en individuos calificados como escépticos y de abierto rechazo a lo sobrenatural. La hipótesis subyacente es pues que, primero, la agentividad tiene un carácter conceptualmente sobrenaturalista, y segundo, que es un mecanismo puramente reflejo soportado por conectividad estructural regulada genéticamente. Valga la contradicción semántica, la sobrenaturalidad de la agencia es natural.4

Dando por sentado que no necesariamente el tamaño sino sobretodo la efectividad del cerebro humano es lo que determina nuestras altas capacidades cognitivas (p.ej. lenguaje) e inteligencia, es así fundamental la estructura del conectoma respecto al FT, por un lado, en tanto sabemos que, si precisamente las ballenas tienen cerebros más grandes (o que los elefantes tienen un córtex más grueso) que el nuestro, en cambio el cerebro humano gana en cuanto a cantidad total de neuronas y el modo más eficiente en que se envía la información entre éstas (ibid. pp. 136-137). Dos factores neuro-evolutivos aparecen pues como necesarios determinantes al respecto: la globularización parietal/temporal y la configuración del conectoma. Y al menos dos genes, microcefalin y ASPM, se han identificado como candidatos de la regulación de ello debido a que, una mutación del primero causa reducción del tamaño del cerebro (comparable al de los primeros homínidos) y retardo mental, mientras que el segundo también se asocia a la microcefalia, aparentemente asociado a la proliferación/diferenciación neural (ibid. p. 136). Es por tanto probable que estos genes, aunque seguramente no sean los únicos, regulen el conectoma y así las capacidades funcionales del FT. Adicionalmente, hay evidencia de presión selectiva sobre estos genes en tiempos tan recientes como entre 37,000 y 5,800 años, así «existe tentativo apoyo vinculando discretos genes candidatos, el tamaño cerebral, y el desarrollo cronológico de las habilidades cognitivas asociadas con los logros humanos relativamente modernos sobre el curso de la historia evolutiva» (ibid. p. 136).
Este modelo de la biología de la inteligencia humana de Jung & Haier (2007), es altamente consistente con el modelo propuesto en el artículo «EL VIAJE MENTAL EN EL TIEMPO…» (y antes en «NEUROCIENCIA DE LA RELIGIÓN» «VI» y «VII») como el sustrato neural de la cognición agentivo-mágico-religiosa, en buena parte conformado por el MTT, que se fundamenta directamente en un ‘sistema temporo-parieto-frontal’ (de acuerdo al sentido direccional del procesamiento de la información) del hemisferio derecho del cerebro como contrapuesto y paralelo al sistema izquierdo enfocado en el lenguaje (Gharabaghi et al. 2009; Karnath 2009). Como muestran Jung & Haier (2007), la inteligencia humana está, precisamente, lateralizada hacia el hemisferio izquierdo. Ahora bien, dado que se discute si el MTT es una facultad exclusivamente humana o no, como fuere, por un lado se ha sugerido como causal neurocognitivo de la religión (Suddendorf & Corballis 1997 resumen, PDF; Bulbulia 2009 aquí o aquí), mientras que por otro, podría también vérsele como indisociable del desarrollo de la inteligencia humana, tal y como se piensa así del lenguaje (Suddendorf & Corballis 1997; Suddendorf, Addis & Corballis 2009; Corballis 2019). De hecho, en el modelo de Jung & Haier (2007) lenguaje e inteligencia están directamente asociados. Así, y de acuerdo al concepto vertido arriba, no parece que pudiera definirse la inteligencia humana sin considerarse vinculada al MTT. No podría, estimo, no ser vista la relación íntima entre la capacidad visuoespacial (básicamente del lóbulo parietal), el lenguaje (lóbulos temporal, parietal y frontal) y el MTT como «el zumo» que llamamos ‘inteligencia humana’, sin a su vez contemplar la fundamental importancia del modo en que se conectan estos lóbulos cerebrales para el efecto y la influencia del genoma en esto.
Conclusión
- Se propone el FT (eje fronto-parietal) como sustrato del «espacio externo como agente intencional»;
- se ha mostrado que la región temporoparietal soporta de manera específica (¿modular?) la espiritualidad o autotrascendencia (Urgesi et al. 2010);
- vemos que la arquitectura y funcionalidad neural involucrada (tanto la estructura y regionalización del córtex como la materia blanca que lo interconecta) están regulados genéticamente;
- por lo tanto se sugiere que estos «canales» cognitivos (‘espectro agentivo-sobrenaturalista’ o ‘agentividad mágica’) que propician y/o facilitan el aprendizaje de la religión, están de modo notable determinados neuro-genéticamente;
- empero, la religión no es un asunto biológico.
NOTAS:
1. Recientemente, Boyer discute sobre el término ‘religión’, que, en tanto refiere a un fenómeno cultural reciente y aún visto en términos de religión occidental (p.ej. las diversas religiones definidas de modo cristianocéntrico), además de entendida como idea/conducta adaptativa por un sector académico (una supuesta base genética para un sistema entero de diversos aspectos desde neurobiológicos hasta puramente culturales pero que servirían exclusivamente a la religión), es un término vacío científicamente. Ver en Aletheia.
SOBRE GENÉTICA, NEUROCOGNICIÓN, PENSAMIENTO MÁGICO Y RELIGIÓN (PARTE 2) (b)
Resumen
Hasta la fecha hay dos aspectos asociados a los rasgos de la personalidad, que a su vez están vinculados a la religión, y para los que un número de estudios arrojan indicios de asociación con material genético, como anotado previamente en «HOLISMO, BIOLOGÍA Y EL ORIGEN DE LA RELIGIÓN»: nos referimos a la inclinación hacia la ‘religiosidad’, y hacia el ‘pensamiento mágico’. Ciertamente, ambos aspectos pueden estar estructurados directamente por o al menos estar asociados a una dimensión de la personalidad comúnmente conocida como ‘espiritualidad’, es decir, un complejo de sensaciones, ideas y conductas que tienden a integrar el yo al universo como un todo: autotrascendencia (Urgesi et al. 2010 p. 310). Está claro que es un término innegablemente complejo en su(s) significado(s), extensible(s) hasta sustraerlo de cualquier interpretación doctrinaria religiosa (p.ej. Fossion 2009, sobre Comte-Sponville y su «spiritualité sans Dieu»); mientras que no está claro si esta separación implica negar cualquier tipo de connotación de trascendencia (respecto a lo que fuere). Así pues, aunque la espiritualidad o autotrascendencia y el pensamiento mágico están vinculados a lo que llamamos ‘religión’, estos no necesariamente se identifican con ella, sino que se trata de aspectos parcialmente independientes (p.ej. Urgesi et al. 2010 p. 316), sobre todo en lo que respecta al pensamiento mágico. En cualquiera de ambos aspectos es esperable encontrar material genético asociado en tanto que en ellos subyace como veremos un número de estructuras neuroanatómicas, funciones y procesos que se sabe están más o menos bajo influencia genética (y medioambiental). Pero sobre esto mismo es necesario hacer otras aclaraciones primero. La espiritualidad o autotrascendencia, por implícitamente tratarse de una «extensión» del yo hacia el mundo externo — una «conexión» causal en el cosmos que involucra al yo de manera más o menos central, no se trata sino de una sensación, idea y/o creencia que podemos conceptuar como «mente-sobre-materia», lo que significa que esto en sí mismo es una manifestación del pensamiento mágico. Similarmente, la religión y la religiosidad también derivan de él, así como muchas otras ideas, creencias y conductas que sencillamente impregnan, cuando no ciertamente estructuran, la entera sociocultura (cual fuere). Lo que se hará en esta segunda parte es descubrirlo como en realidad un modo de funcionamiento mental por defecto, en lugar de una invención cultural.
1.1. RELIGIOSIDAD
La religiosidad (R) se trata de un auténtico sistema o complejo de elementos conductuales, afectivos, culturales, comunales e individuales sobre las convicciones, el grado de compromiso y la participación enfocadas en las creencias religiosas. Tratando de dar una definición objetiva, para así facilitar la medición y el testeo empírico del grado de R en las personas, se ha postulado el concepto de ‘dimensiones de R’, y diversos estudiosos básicamente han identificando un número de dimensiones que abarcan más o menos tales aspectos (Holdcroft 2006). Dimensiones como devoción, afiliación, conocimiento doctrinal, creencia, asistencia a un centro de culto y espiritualidad han sido propuestas, habiendo correlaciones p.ej. entre el aspecto comunal y de integración social con la pertenencia denominacional (ibid. p. 91). La discusión de tales dimensiones y la diversidad de propuestas teóricas (aunque de hecho muy similares entre sí) responden al simple hecho de que, como se observa, una persona puede ser muy creyente pero no asistir a un culto. La evidencia de heredabilidad aquí no es uniforme, siendo que los estudios con gemelos y mellizos sobre la afiliación religiosa «han demostrado que la variación en este rasgo es de origen casi completamente medioambiental», mientras que «la frecuencia de la asistencia a la iglesia [...] parece estar genéticamente influenciada» (Bouchard & McGue 2003 p. 29). Ya que una relación entre herencia genética y R es sugerida por estudios que se siguen acumulando (p.ej. recientemente Button et al. 2011), habría que suponer que lo heredable sean precisamente estas dimensiones como parte, probablemente, de los rasgos de la personalidad, p.ej. si una marcada pertenencia denominacional se vinculase a la extraversión; o como sugiere un estudio mostrando numerosas actitudes conductuales con relevante índice de heredabilidad (tan variadas como la actitud hacia el atletismo, a jugar bingo, al aborto, al liderazgo, a la religión organizada, etc.), mediadas a su vez por rasgos de la personalidad también con índice de heredabilidad (Olson et al. 2001).
Entre tanto, la R parece gozar de una recurrente valoración positiva, seguramente tratándose tal valoración de un asunto popular y/o de una herencia cultural, yendo a la par de la literatura médica que la asocia p.ej. con el mejoramiento anímico o de la salud (p.ej. Fehring et al. 1997; Hackney & Sanders 2003; Koenig 2004). Sin embargo, igualmente hay resultados negativos, por lo que «la evidencia de una asociación entre religión, espiritualidad, y salud es débil e inconsistente», y entonces «es prematuro promover la fe y la religión como un tratamiento médico complementario» (Sloan et al. 1999; ver también: Sloan & Bagiella 2004; Sloan 2008). De hecho la R se asocia también al aumento de la ansiedad y el estrés (porque es sobre una divinidad tan benevolente como castigadora: Wilt et al. 2015; O'Brien et al. 2018), se asocia a la inseguridad existencial (Barber 2011), a la desigualdad económica —e incluso a ser probablemente un medio con el cual los ricos manipulan a los pobres (Solt et al. 2011), o a la misoginia, la homofobia, el conservadurismo, y el racismo (Glick et al. 2002; McPherson 2019; Whitley 2009; Rowatt et al. 2009; Whitehead & Perry 2019). Con todo esto, estamos lejos de permitir conclusiones positivas y más bien encontramos mayores efectos negativos de la R más allá de la medicina (y que resultan en daños a la salud pública), o bien los datos son simplemente ambiguos (Thuné-Boyle et al. 2006). Sin embargo tampoco es coherente una concepción patológica de la R (Bergin 1983). Finalmente, si bien la hipótesis adaptacionista de la religión toma como «evidencia fuerte» las altas tasas de fertilidad entre las personas con mayor grado de afiliación religiosa (Blume 2009; lo que, según el autor, es evidencia de una selección genética de la R), se tiene por otro lado que el éxito reproductivo correlaciona con ciertos rasgos de la personalidad: neuroticismo femenino y extraversión masculina (Alvergne et al. 2010). Y hay, por supuesto, una asociación entre extraversión y R (p.ej. Francis et al. 2010; Shachar et al. 2011 p. 9; Colledani 2017).
Todo esto sugiere más bien que no habría «genes de la R» (ni de la religión) sino, en todo caso, de ciertos rasgos que se asocian o median las dimensiones de la R, tanto como de otros aspectos conductuales, p.ej. vemos, asociados a la reproducción. Así pues, hay también robusta evidencia que vincula la R a factores medioambientales socioculturales. Un reciente estudio comparativo de 645 rituales religiosos de 74 culturas indica que ellos están vinculados al tamaño del grupo y su estructura, y que las religiones «doctrinales» (p.ej. basadas en libros sagrados) con alta frecuencia ritual (en contraste con las religiones «imaginistas», no centralizadas y con baja frecuencia ritual), van ligadas históricamente a la llegada de la agricultura y la posterior aparición de las primeras civilizaciones a gran escala (Atkinson & Whitehouse 2011). La R está ampliamente correlacionada con factores socio-económicos, al punto de que su fluctuación, o sea si los individuos son más religiosos o si de hecho abandonan las creencias religiosas, parece depender de la eficiencia del bienestar social como proyecto gubernamental (Gill & Lundsgaarde 2004). El asunto va más lejos aún: eso a su vez correlaciona con el factor ‘éxito de la democracia’ y del ‘mercado liberal’ ciertamente restringido a los países del primer mundo («prósperas democracias desarrolladas»), y así «las naciones menos teístas son usualmente las menos disfuncionales» democrático-económicamente (Paul 2005 p. 7). Estos y otros estudios sociológicos conforman un amplio cuerpo de evidencia de que la R depende fuertemente de factores no estricta o no reductiblemente neurobiológicos. Sin embargo, se trata de un asunto obviamente muy complejo donde necesariamente las encuestas y los censos, y sobre todo esa evidente perspectiva político-económica (naciones «prósperas» vs. «disfuncionales» así definidas desde una filosofía de mercado), deben dialogar con la ciencia cognitiva y la antropología para elaborar un marco explicativo completo de este fenómeno religioso/social/económico, porque, como último ejemplo, hay evidencia de que la preferencia por el creacionismo religioso (vs. el evolucionismo científico) depende de mecanismos cognitivos respecto a la falta de control (Rutjens et al. 2010), en lo que otros estudios han mostrado que lo que llamo ‘espectro agentivo-sobrenaturalista’ es un regulador natural (de la sensación de falta de control).1 Una vez más, lo que decide las inclinaciones religiosas/teístas no son genes expresamente dedicados a eso.
1.2. PENSAMIENTO MÁGICO
El pensamiento mágico (PM) «primariamente compromete una tendencia a asumir significados ocultos en las configuraciones aleatorias y a insistir en una determinación causal de las coincidencias» (Mohr et al. 2003 p. 169; ver «NEUROCIENCIA DE LA RELIGIÓN (II): DOPAMINA, PENSAMIENTO MÁGICO Y RELIGIÓN»). «Generalmente llamamos a algo mágico o supersticioso si ello implica agencia humana (como diferente de la religión), e invoca causas inconsistentes con los actuales conocimientos, mediante los pertinentes “expertos” (p.ej. los científicos occidentales), de cómo funciona el mundo» (Wilson & Keil 2001 p. 503 —libro completo disponible aquí). De hecho, una asunción intrínseca e implícita del PM, y lo que necesariamente lo distingue del pensamiento científico y el realismo fisicalista, es el de «mente-sobre-materia» (y por extensión «mente-sobre-mente»): «los objetos físicos en el final receptivo de un proceso comunicativo tienen algún tipo de consciencia» (Subbotsky 2010 p. 9 —libro completo disponible aquí). Es decir, cuando se percibe o se idea (consciente o inconscientemente) que dos eventos u objetos están causalmente relacionados de un modo tal que excede el fisicalismo científico, tal causalidad implica fundamentalmente que esos objetos o eventos ‘saben’ y aún ‘tienen la intención de’ comunicarse.
A través del desarrollo del individuo, a partir de aprox. los 5 años de edad, el PM y las creencias mágicas resultan más o menos decaer, coincidiendo con la adquisición de explicaciones fisicalistas de los objetos y el mundo, pero no desaparecen (p.ej. los niños pueden negar lo mágico-anticientífico aludiendo a trucos ilusionistas, pero cuando son enfrentados a trucos que no pueden explicar endorsan PM, aún cuando se les explica en qué consiste el truco). Acercándose la adolescencia y con el asentamiento de la educación científica, el PM parece desaparecer, pero a la vez puede manifestarse y aún reafirmarse bajo nuevas formas debido a la presión cultural misma (p.ej. los medios incentivando el PM, o la religión), lo que desemboca en una divergencia de las creencias mágicas en «no institucionalizadas» (condenadas por la religión doctrinal), e «institucionalizadas» (la religión en sí misma). Finalmente, en la adultez el PM «persiste sin obstáculos y desarrolla nuevas formas, como disfrutar de las tendencias en el arte que implican objetos y eventos mágicos (el surrealismo en la pintura, el realismo mágico en la ficción, las formas avanzadas de la música) o, en el lado negativo, estimular artificialmente el compromiso con la realidad mágica a través del uso de drogas alucinógenas» (ibid. pp. 164-165). Entre tanto, hay que aclarar que el PM es un estilo cognitivo, una clase de inferencia intuitiva más bien que una creencia:
«Una importante distinción hay que hacer entre el pensamiento mágico y las creencias mágicas. En contraste con el pensamiento mágico, el cual confina los carácteres y eventos mágicos al dominio de la imaginación, las creencias mágicas implican que los carácteres o eventos mágicos existen en el mundo físico real.» (ibid. p. 7).Con el PM no hay tal enfoque de esa misma naturaleza socio-económica como ocurre con la religiosidad (R). A saber el PM, en sus variadas formas desde la superstición, pasando por el amplio campo de «lo paranormal», hasta las representaciones post-vida (agencia de la muerte), ha sido observado en individuos con educación avanzada o específicamente científica y aún entre ateos y personas declaradamente escépticas en la post-vida (p.ej. Bering 2010 p. 2 —abstracto, artículo completo; Bering & Heywood 2010, estudio no publicado del SPSP 2010 Annual Meeting Programm pp. 32-33, abstracto aquí; ver también Nelson et al. 2020).2 Puntuar alto en análisis lógico o racionalidad tampoco es independiente del PM ni lo excluye, ya que de hecho en un mismo individuo co-existen dos modos paralelos de procesamiento cognitivo: intuición y racionalidad —entre tanto, el PM tampoco implica poseer poca inteligencia (Hood 2009 pp. 245-246 —libro completo disponible aquí). (De hecho, ya anteriormente hemos tratado el tema de la intuición: no necesariamente significa «irracionalidad» y es un procesamiento perceptual/cognitivo/emocional primario, inmediato y altamente adaptativo —ver «LA INTUICIÓN NO ES IRRACIONAL».)
El PM abarca extensos aspectos bastante mundanos, habiendo en principio grupos sociales que son, «por naturaleza o necesidad, particularmente supersticiosos» como los deportistas, los jugadores o los estudiantes (Vyse 2000 p. 26). Tenemos p.ej. las ideas sobre la asociación comida/salud (Lindeman et al. 2000), los conceptos sobre los exitosos hombres de negocios y los políticos importantes (Schultheiss & Brunstein 2010 p. 413: «para sus seguidores, tales líderes pueden parecer tener poderes casi sobrenaturales, sobrehumanos, o mágicos»), el éxito en la audiencia de los gurús del manejo organizacional (Clark & Salaman 1996), la fidelidad a una marca/producto del mercado (Muñiz & Jensen 2005; incluso se teoriza que el culto a la marca puede sustituir a la religión: Shachar et al. 2011), la percepción de influencia personal en una competencia deportiva, hasta la inducción engañosa de causalidad mágica en estudiantes de Harvard que aún no creyendo en muñecos vudú, actúan muy sutilmente como si ellos mismos sospecharan de realmente inducir por ese medio malestar físico en un individuo (Pronin et al. 2006). No es exagerado afirmar que el PM está en todos lados, en cualquier marco sociocultural, geográfico, de edad, en la educación científica o en el entretenimiento (Shweder 1977; Diaconis 1986; Harris 1997; Rosengren & Johnson 2000; Elkan 2001; Schultheiss & Brunstein 2010; Passanisi et al. 2017; Kiken 2019).
Las inferencias sobre la personalidad de la gente también caen en esta categoría, en tanto que, «en su mayor parte, los rasgos de la personalidad no se pueden descubrir en el comportamiento, sino que son las creaciones de la mente mágica» (Shweder 1977). Nótese que esto último implica que la ToM, de lejos intrínsecamente más probabilística y especulativa que cualquier hipótesis medianamente objetiva sobre la conducta, no es sino PM, lo que va en apoyo conceptual de que, en lo que insisto, el ser humano pueda definirse como un sobrenaturalista nato.3 «El pensamiento mágico es una característica universal del pensamiento cotidiano» (ibid.), en contraste con R, luciendo como un latente y subyacente rasgo cognitivo identificable en prácticamente cualquier aspecto psicológico, social y cultural del ser humano. De hecho, «una forma controlada de pensamiento mágico —en el modo de “hipótesis de trabajo”— es un básico ingrediente del progreso científico» (Diaconis 1986 p. 2); mientras que «es a menudo asumido que el pensamiento científico desplaza al pensamiento mágico, tanto histórica como ontogenéticamente, pero la tendencia a invocar poderes ocultos causales cabalga entre ambos modos de pensamiento» (Harris 1997). Toda esta enorme cantidad de evidencia acumulada refuta contundentemente la tradicional idea del ‘reemplazo lineal PM → religión → ciencia’, ya que demostradamente, el PM simplemente persiste, a pesar de la educación científica (Rosengren & Johnson 2000 p. 16).
Si el pensamiento mágico (PM) es verdaderamente universal, o mejor dicho, si es un default cognitivo, en contraste con la religiosidad (R), entonces debe poder hallarse en principio, evidencia conductual de que no solo existe en cualquier cultura, cosa que ya se sabe, sino de que efectivamente persiste en la mente de los individuos (a un nivel inconsciente, aunque el individuo verbal y conscientemente lo niegue) con adquisición de conocimiento científico y a pesar de ello (asumiendo pues que el PM y el pensamiento científico son mutuamente excluyentes). Para empezar, varios estudios aplicando cierta metodología muestran que las respuestas verbales de un individuo en el laboratorio pueden diferir de su comportamiento cotidiano real. «Como se ha demostrado en muchos estudios, las respuestas conductuales en condiciones que pueden implicar ‘alto costo’ (es decir, tienen consecuencias importantes para las necesidades primarias de los participantes) reflejan con mayor precisión las ‘creencias reales’ de los participantes de lo que lo hacen las respuestas verbales de los participantes, que son sensibles a factores como las expectativas sociales, las fallas de memoria, y el limitado conocimiento de las propias necesidades y creencias (Thomas, 1971; Wilson y Nisbett 1978)» (Subbotsky & Quinteros 2002 p. 520). Estos autores aplicaron tales condiciones a sendos estudios realizados con habitantes rurales mexicanos (en México) y estudiantes universitarios ingleses (en Inglaterra, realizado previamente), sobre sus reacciones ante un evento inusual, para medir sus adhesiones a las explicaciones causales científicas o mágicas; y luego compararon los resultados obtenidos con ambos grupos. El alto costo se refiere a una condición experimental en la que p.ej. el individuo sepa que puede ser contagiado por una enfermedad grave como el SIDA, lo cual previamente se ha mostrado que incrementa los juicios de PM sobre la enfermedad misma (ibid. p. 539). En este caso, el bajo costo consistió en estropear accidentalmente un documento importante al ponerlo dentro de una caja que repentinamente destruía lo que estaba en su interior (mediante un dispositivo que era visible para el «contexto científico», y estaba oculto para el «contexto mágico»); mientras el alto costo implicaba meter la propia mano en la caja.
«Si la hipótesis del ‘efecto profundo’ de la racionalidad científica en un individuo es verdad, entonces el grado de credulidad de los participantes ingleses hacia las explicaciones mágicas debe ser significativamente menor que la de los participantes mexicanos. Entre tanto, si bajo las mismas condiciones los participantes ingleses y mexicanos mostraran credulidad hacia la magia en la misma medida, esto significaría que, en un cierto nivel fundamental, la mente de un individuo occidental queda relativamente poco afectada por la racionalidad científica.» (ibid. p. 525)Tal como predicho, la comparación de los resultados mostró que los ingleses y los mexicanos fueron por igual conductualmente mágicos bajo las mismas condiciones de alto costo; y sólo a un bajo costo los ingleses fueron más racionalistas científicos que los mexicanos. Como los autores discuten, la corriente cultural occidental predominante entre los estudiantes ingleses es la racional-científica (p.ej. un mayor contacto con la tecnología en contraste con los habitantes rurales mexicanos, que haría a los ingleses más escépticos del «misterioso» comportamiento de la caja), pero está prácticamente ausente en una comunidad rural mexicana (considerada incluso no-occidental), y es lo que evidentemente modula el rechazo o la preferencia del PM respectivamente. Pero, lo aquí descubierto es que un condicionante ansiogénico (el potencial perjuicio propio) puede «desviar» al individuo de sus creencias científicas adquiridas (ibid. p. 540). En otras palabras, sumado esto al amplio data de las diversas manifestaciones de PM en aspectos y entornos culturales modernos (científicos y tecnológicos), se puede sugerir con firmeza que el PM tiene un fundamento biológico, y que la cultura solo lo modula superficialmente, bien asentándolo en forma de creencias y prácticas explícitas, o bien disminuyendo su influencia en la elaboración de creencias, pero, por lo visto, sin lograr eliminarlo a un nivel cognitivamente profundo. Este argumento por supuesto no se funda en el único estudio que se acaba de comentar, sino que hay una considerable cantidad de resultados similares con niños y adultos (con relativa educación científica), que demuestran que el PM persiste (Albas & Albas 1989; Rosengren & Hickling 1994, 2000; Rozin et al. 1990; Woolley 1997; Subbostky 1997, 2010, 2014; Risen 2016; Garrett & Corte 2017; Wilson 2018).
En artículos previos se ha buscado, como parte de la tarea de este sitio, definir la naturaleza humana analizando específicos rasgos cognitivos como productos de la hominización y en particular del desarrollo de específicas partes y funciones del cerebro humano (p.ej. agencia, ToM, mental time travel —MTT), que producen elevados y complejos órdenes de metarrepresentación mental (Pyysiäinen 2009 p. 19: «yo sé que Juan quiere que María entienda que Pedro cree que Isabel lo ama») que no se equiparan a nada hasta ahora visto en nuestros parientes primates más cercanos, y mucho menos en el resto de mamíferos. Aunque parezca complicada o confusa esta tarea de integrar todo ello en una pretendida teoría de la naturaleza humana, al menos un hecho sugerente es que neuroanatómicamente en efecto hay una fuerte integración identificada cognitivo-conceptualmente con el término agentividad.4 Así, «agentes sobrenaturales, fantasmas, y pensamiento mágico [...] todos pueden ser explicados por el mecanismo general hiperactivo de detección de agencia»5 (Beit-Hallahmi en Bering 2006 p. 464). En realidad, sería más adecuado denominar a esto AGENTIVIDAD MÁGICA, esto es, la atribución de agencia o intención a prácticamente todo, integrada a la capacidad de viajar imaginariamente en el tiempo/espacio, posibilitando así escenarios contrafactuales, con alcances profundamente emocionales y altamente abstractivos, conformando todo junto una función permanente y automática (como en efecto se vió en la primera parte de este artículo). El PM implícitamente forma parte central de esto, y por ello su universalidad y carácter generalista. Como aspecto manifiesto de agentividad, tiene pues toda la apariencia de un default cognitivo:
«El pensamiento mágico ha sido encontrado entre gente viviendo en culturas tribales (p.ej. Golden 1977), personas experimentando psicosis en esquizofrenia o desorden bipolar (p.ej. Thalbourne & French 1995), y jóvenes infantes que aún han aprendido los principios de la ciencia (Piaget 1929; Woolley 1997). Lo que probablemente sorprenda a la gente es saber que destellos de pensamiento mágico aparecen incluso en personas y circunstancias ordinarias cuando los eventos conspiran para promoverlo.» (Pronin et al. 2006 p. 218)Decía líneas atrás que es falsa la idea del reemplazo del PM con la religión y luego con la ciencia: el PM no solo persiste aún con o sin educación científica sino que existe también con o sin religión. Tal como con la autotrascendencia, PM y R co-dependen: en tanto que la hiperagentividad ha sido invocada como estructura importante para ambos «es justo decir entonces, que el pensamiento mágico es un elemento clave de la creencia religiosa» (Markle 2010 p. 22), pero similarmente, puede haber PM sin R. P.ej. Tobacyk & Pirttilä-Backman (1992) hallaron que, comparando a estudiantes de EE.UU. y Finlandia, los estadounidenses fueron más creyentes sobre religión tradicional, superstición, brujería, y vida extraterrestre, manifestando también mayor anomia, mientras que los finlandeses mostraron más preocupación sobre la muerte. Más interesante aún fue que «las creencias paranormales mostraron relaciones más fuertes con las medidas de ajuste de la personalidad en los finlandeses que en los estadounidenses», en tanto que estos resultados «son consistentes con la noción de que la secularización ha avanzado más en Finlandia que en EE.UU.» O sea que la secularización efectivamente deprime la convicción religiosa (en lo que es muy probable que estén implicados los factores socio-económicos antes comentados), pero no al PM.
Previamente, Tobacyk & Wilkinson (1990), mientras habían encontrado notorias relaciones entre PM, espiritualidad, psi, precognición, superstición y brujería, también encontraron que «las creencias religiosas mostraron una significante relación inversa con el pensamiento mágico en hombres solamente». Seguramente esto tiene que ver también, en cuanto a mero aprendizaje cultural y no alguna predisposición neurocognitiva, con que ciertos esquemas religiosos p.ej. cristianismo, que es la religión que típicamente manifiestan los individuos en estos trabajos referidos efectivamente regulen, virando aparentemente hacia la supresión (recordemos la evidencia con ingleses y mexicanos), las creencias mágicas de las personas. En efecto, las estadísticas, en EE.UU., indican que solo 8% de personas religiosas creen en precognición, Atlántida, Piegrande, casas embrujadas y la comunicación con los muertos, contrastando con un 31% de personas irreligiosas creyendo en los mismos tópicos.6 Al fin y al cabo, aunque las creencias mágicas puedan ser «encubiertas» con ciencia o bien con religión, el PM demostrablemente subsiste de dos maneras: inconscientemente o como un continuum (más o menos consciente) implícito de las creencias religiosas mismas. Una robusta observación en consonancia, finalmente, con que las intuiciones primarias en general son suprimidas pero no suplantadas por la ciencia, coexistiendo cognitivamente (Shtulman & Valcarcel 2012), por lo que una compresión de p.ej. la ilusión de causalidad debería servir de herramienta para la educación científica (Matute et al. 2015).
2. Pensamiento mágico: ansiedad y control
Se han desarrollado diversas explicaciones sobre el PM. De acuerdo con Pronin et al. (2006 p. 219), se trataría de una «transferencia de la “esencia” entre los objetos y como si el traslado de la esencia de tales creara una conexión entre las dos entidades, así, por ejemplo, haciéndonos indeseable llevar una prenda lavada una vez usada por un nazi (Rozin et al. 1986; Rozin & Nemeroff 2002)». O bien «podría ser un vestigio de la infancia cuando las concepciones científicas de causalidad son menos entendidas (y menos arraigadas culturalmente)»; aunque los adultos, al igual que los niños, puedan verbalmente negar endorsar PM, su comportamiento sigue evidenciándolo. «También puede ser el resultado de errores cognitivos comunes que implican el uso de atajos mentales o heurística», implicando p.ej. la inferencia de que la semejanza de dos eventos significa que uno causa el otro, así permitiendo no solo creencias mágicas sino también inferencias racionales, respectivamente p.ej. «el sol es de color amarillo y el cielo es azul, por lo que juntos hacen la hierba verde»/«la lata de pintura es de color verde y el charco mojado en el suelo es de color verde, por lo que el charco debe ser de la pintura». Esta perspectiva del PM como error es seguida por Lindeman & Aarnio (2008), quienes profundizando más en el desarrollo psicológico infantil y el uso de los «núcleos de conocimiento» (grupos de conceptos, inferencias y expectativas intuitivas e innatas, asumidos como ‘módulos’ cognitivos, sobre categorías más o menos específicas de objetos y eventos: ‘folk psychology’, ‘folk biology’, ‘folk physics’, etc.),7 proponen el PM como la «confusión» o traslapado de tales núcleos resultante de la maduración cognitiva aprox. a partir de los 3-4 años de edad. Entre tanto, «viniendo más desde una perspectiva motivacional, otra explicación para el pensamiento mágico sugiere que se produce, sobre todo en tiempos de incertidumbre o estrés, para servir a una necesidad motivacional de control» (Pronin et al. 2006 p. 219). Creo que estas aproximaciones, lejos de ser excluyentes entre sí, pueden más bien integrarse todas en torno a tres aspectos:
- El procesamiento primario de la información sensorial, de carácter perceptual-categorial y modular (p.ej. la «esencia» de un objeto puede no ser otra cosa que alguna folk), a un nivel neural básico, como p.ej. el sugerido ‘eje de la agentividad’ temporo←→parietal, en «EL VIAJE MENTAL EN EL TIEMPO (‘MENTAL TIME TRAVEL’) Y LA COGNICIÓN RELIGIOSA» Fig 3. Es de capital importancia que haya evidencia de que categorizaciones perceptuales como p.ej. «viviente»/«no viviente» «están innatamente dispuestas para manejar la información» (Mahon et al. 2009; Farah & Rabinowitz 2003); o el reconocimiento facial, otro proceso perceptual intrínseco de la agencia, sea heredable (Zhu et al. 2010), y en consecuencia responda a un fenotipo (Sarpal et al. 2008). De hecho se ha sugerido antes el reconocimiento facial/corporal/agentivo como un modelo neurocognitivo de la ‘contraintuición religiosa’ (ver «NEUROCIENCIA DE LA RELIGIÓN (V): CUANDO LAS INTENCIONES DEL ENTORNO TIENEN ROSTRO»), que en sí misma se trata de PM, como dicho en ese artículo referido: una base neurocognitiva que de modo natural, automático o inconscientemente, genera percepciones de intención e imágenes de corporeidad ante objetos inanimados o circunstancias complejas.
- La producción de representaciones que pueden adquirir forma narrativa con carácter lógico y crediticio (p.ej. las ideaciones resultantes del traslapado de folks y las inferencias intuitivas y racionales) a un nivel neural más complejo (p.ej. la ‘red por defecto’ temporo←→parieto←→frontal) y susceptible de eco-modulación (influencia sociocultural). El hemisferio izquierdo del cerebro se ha conceptualizado como un «intérprete» del derecho, tratándose de un proceso ciertamente prefrontal, mientras que las cortezas prefrontodorsolateral/prefrontomedial/inferoparietal derechas están más activas a la hora de predecir patrones (ver «EL HEMISFERIO IZQUIERDO: PATRONES, HIPÓTESIS E IDEAS DELIRANTES»). Ahora bien, todo esto da cuerpo a una propuesta propia de modelo explicativo de la cognición religiosa (ver aquí), a su vez consistente con el modelo de desarrollo jerárquico sobre un mismo núcleo cognitivo ‘[movimiento biológico→]agencia→ToM’ que propone Horst (2010),8 donde tanto el dualismo innato (Bloom 2004; 2007) y el animismo (Guthrie 1980; 1993; 2002) resultan de la natural temprana disociación de folks, así evidenciada psicológicamente (aunque, neuralmente no están necesariamente disociadas: ver aquí), mientras que la contraintuitividad resulta del traslapado de folks (Boyer 2001) (aunque, en realidad este traslapado mismo emerge naturalmente en el neurodesarrollo: ver aquí). Así, dualismo y contraintuitividad están similarmente integrados, sin excluirse, formando parte de un desarrollo cognitivo jerárquico, con ciertos aspectos neurales compartidos.
- 1 y 2 pueden obedecer juntos a una promoción de la sensación de control u otras ‘motivaciones’ (de apariencia instintiva), que es una regulación de carácter ansiolítico y del estrés (porque la interacción con el medio —natural/social— puede ser ansiogénica y estresora), de nivel neuro-endocrino de funcionamiento, aunque aparentemente dependiendo de qué tan ansiogénico pueda ser percibido un estímulo según la circunstancia y el contexto. «El aumento repentino de la carga de ansiedad podría ejercer como un disparador que temporalmente cambia la forma en que la mente humana procesa la información de los métodos estándar hacia el pensamiento mágico» (Markle 2010 p. 28). «Si lo mágico es el resultado de una confusión primitiva, un avance del desarrollo (en el caso del contagio), la arquitectura de dominio general del sistema humano del procesamiento de la información, una extensión natural de la experiencia corporal, o modos preprogramados o pre-adaptados de pensamiento de dominio específico — claramente suele ser útil [...] mapea las contingencias del mundo real [...] también provee explicaciones satisfactorias [...] El pensamiento mágico es a veces adaptativo y otras problemático, pero casi siempre es una fuerza a tener en cuenta» (Rosengren & Johnson 2000 p. 29; cf. Subbotsky 2010 pp. 170-173).
Así, agregando aquí que el núcleo de tales componentes, la atribución de agencia (p.ej. Horst 2010), implica la sensación de control, que, además, «los sentimientos de frustración son consecuencia de una sensación de falta de control» (Barrett & Johnson 2003 p. 23), y que tanto la falta de control y la frustración son estresores (cf. Almeida et al. 2002) o incluso pueden verse como un tipo de estrés (Weiten et al. 2008 p. 75), tenemos pues dichos ejemplos como pareidolia, correlaciones ilusorias y percepción de conspiraciones en situaciones de falta de control (Whitson & Galinsky 2008), el incremento del PM (pensamiento mágico), la superstición y la causalidad bajo condiciones de estrés (Keinan 2002; Keinan & Tal 2005; Lasikiewicz & Teo 2018), y ver los desastres naturales como manifestaciones de intencionalidad (Bering & Parker 2006 pp. 253-254). Incluso hay evidencia de que la superstición mejora la performance motora y cognitiva (Damisch 2008; Damisch et al. 2010; sin embargo ver Calin-Jageman & Caldwell 2014). Pronin et al. citan algunos estudios que acabamos de referir, vertidos también en la Parte 1 de este artículo y en otro previamente, siendo otros estudios necesarios de agregar para terminar de sentar el vínculo PM/estrés/ansiedad. Tenemos el PM documentado en personas habitando entornos, en períodos entre guerras, con alto nivel de desempleo e inestabilidad política; en policías con trabajos peligrosos; infectados de VIH con expectativas negativas sobre su propia salud (Taylor et al. 2000: «las investigaciones han revelado que aún las creencias irrealistas optimistas sobre el futuro pueden ser protectoras de la salud»); o en jugadores de lotería. «Incluso cuando las personas reconocen que el control de los eventos de la vida puede ser imposible de lograr, las creencias mágicas pueden emerger de una motivación por buscar el “significado” de lo que no pueden controlar» (Pronin et al. 2006 p. 219).
Con todo esto, es de esperarse que en una edad avanzada, hacia el final de sus días envueltas en ansiedad y estrés, las personas naturalmente manifiesten PM, implícito en el aumento de religiosidad y/o espiritualidad así observado (Lavretsky 2010). Ciertamente, esto último se asocia a otro asunto del envejecimiento para ambos mostrarnos simultáneamente que, además de lo visto por Lavretsky (2010), el PM en efecto no desaparece como estilo intuitivo sino que «resurge» o es «liberado» cuando se deterioran las funciones inhibitorias (del córtex prefrontal), que es justamente lo observado en pacientes con Alzheimer, en forma de «promiscuidad teleológica» típicamente infantil (Lombrozo et al. 2007), siendo «animistas» (Zaitchik & Solomon 2008: «creemos que esto refleja una disposición innata a ver entidades activas como agentes»), y, evidenciando un específico daño en su folk biology respecto a la «conservación de la especie» (a diferencia de una folk physics ciertamente intacta), consideran que un mapache puede convertirse en una mofeta tan solo por cambiarle el pelaje e introducirle una bolsa de olor apestoso en el cuerpo (Zaitchik & Solomon 2009 pp. 6-7), inferencia típica, nuevamente, del PM infantil. Como Lombrozo et al. (2007 p. 1004) discuten, el Alzheimer no solo devasta la memoria, sino que puede ser mejor caracterizado como un problema inferencial a nivel intuitivo. Entre tanto, el estrés se asocia al Alzheimer involucrando un proceso bioquímico causal que afecta al hipocampo y por tanto la memoria y el aprendizaje (Rissman et al. 2007; Kang et al. 2007; Bolós et al. 2017), lo que es sugerente de que aquel desborde cognitivo-mágico sea parte de un mecanismo de control o amortiguamiento de cara a esta patología. Y esto porque las inferencias basadas en el concocimiento científico adquirido pueden deteriorarse, pero no las inferencias teleológicas, animistas ni contrafactuales, lo que es más evidencia de que el PM es un fenómeno cognitivo profundo, innato, y probablemente, protectivo de los efectos del estrés y sus implicaciones causales en diversos trastornos. Todo aún sabiendo que el PM también ayuda a generar creencias problemáticas y potencialmente dañinas.
Ahora bien, por un lado, en tanto que el PM con todos sus aspectos estructura la religión en general, y que la convicción religiosa compensa la falta de control (Kay et al. 2010) o que la meditación de contenido u origen religioso tiene efectos anti-estrés (p.ej. meditación kundalini: Lazar et al. 2000; yoga: Speca et al. 2000; Eppley & Abrams 1989), emerge una pregunta lógica: «¿es la religión un ansiolítico?» Retomando lo respondido en ese artículo: hay evidencia que sugiere que sí, pero, es debido a que el cerebro de por sí busca minimizar la percepción del error y tiene ciertas predisposiciones [PM ~ agencia, teleologismo, dualismo], que las creencias religiosas pueden ser ansiolíticas. Sabemos además que la interacción social es un estimulante fisiológico/emocional [como veremos luego], lo que sugiere que el solo hecho de compartir creencias puede multiplicar el efecto ansiolítico. En suma, este efecto en realidad no puede limitarse a las creencias religiosas, ni pertenecer exclusivamente a estas, sino que debe funcionar con cualquier tipo de creencias o conductas (Inzlicht et al. 2009) a las que uno se aferre en tanto que, primariamente, nos proporcionen sensación de control. De hecho, se ha mostrado que la afiliación política conservadurista también atenúa la actividad del CCA [córtex cingulado anterior] (ibid.). Como agregó Inzlicht en NewScientist, algo similar ocurriría con los fans devotos a un equipo deportivo. Así pues, la religión ‘en sí misma’ no es un reductor de la ansiedad y el estrés, sino el PM, que ahora sabemos está presente en la política, el deporte o en la economía.
Por otro lado, en el punto (3) sugiero que (1 y 2) obedecen a otras motivaciones además del control: me refiero a la necesidad de pertenencia, grupo y socialización. Ésta tiene un profundo impacto cognitivo/emocional, «de hecho, nuestro sentido del yo deriva de los grupos y categorías a las que pertenecemos, y de muchas maneras la individualidad puede “simplemente” ser una combinación única de distintos grupos y categorías que definen quienes somos. La centralidad permanente de los grupos en la vida humana puede incluso haber tenido un impacto evolutivo en nuestra especie» (Leary & Tangney 2005 p. 462). Entre tanto, sabemos que esto no se trata de una motivación independiente ni disociada de la necesidad control. Ya que control/socialización están vinculados, y que control/PM/religión también lo están, evidentemente socialización/religión deberían entonces estar asociadas: en efecto, la necesidad de pertenencia puede motivar la creencia en Dios (Gebauer & Maio 2011). Asimismo, es de esperarse un vínculo estrés/religión, que aunque ya conocido mediante otras líneas de investigación, no significa que la religión siempre, o necesariamente, es anti-estrés, lo que se explica así: la felicidad y la satisfacción que un individuo siente con su vida, aunque asociada a la religión, está mediada no por la creencia religiosa per se sino por la inclusión en una red social (y así los lazos socio-afectivos involucrados, que sabemos regulan la ansiedad y el estrés) que implica la participación religiosa (Lim & Putnam 2010). Un intrincado sistema socioconductal, neurocognitivo y fisiológico, de flujo funcional bidireccional ↑↓ (de abajo hacia arriba y viceversa en la conexión cuerpo-cerebro) es la base de estas asociaciones, sus causas (p.ej. control ←→ socialización ←→ regulación del estrés) y sus efectos (regulación del estrés ←→ control ←→ PM), con ciertos elementos claves como la oxitocina o la dopamina (ver «¿Por qué la religión hace a la gente más feliz? Pista: no Dios»).
3. Control, recompensa, socialización y regulación del estrés: una aproximación neurobiológica al pensamiento mágico
«El control personal ha sido una de las más pervasivas y duraderas ideas en la investigación y teorización psicológica. Numerosas teorías postulan un importante rol para los constructos de control en la conducta humana tales como autoeficacia (Bandura 1977), causación personal (deCharms 1968), motivación de efectancia (White 1959), control percibido (Thompson 1981), desamparo (Seligman 1975), autocontrol (Baumeister & Heatherton 1996), control conductual percibido (Ajzen 1985), control en el envejecimiento (Baltes & Baltes 1986; Lachman 2006), control primario y secundario (Rothbaum, Weisz & Snyder 1982), control compesatorio (Schulz & Heckhausen 1996), controlabilidad (Heider 1958), y agencia (Deci & Ryan 1987).Nuestro escudriñamiento del PM nos conduce a la ilusión de control, una necesidad natural como poderoso aspecto motivacional que vemos se manifiesta diversamente ideacional y conductualmente. «Uno de los motivos más importantes de la conducta humana es la necesidad de adquirir suficiente competencia para controlar el ambiente. Los teóricos abordan este concepto en formas ligeramente distintas, pero todos coinciden en que el individuo sano necesita estar convencido de su capacidad de influir en el entorno» (Petri & Govern 2006 p. 361). La ilusión de control está íntimamente relacionada a la agencia: «refleja un mecanismo básico que ayuda a las personas a mantener» la segunda (Elk et al. 2015), y proporciona «estimaciones más altas de probabilidad de recompensa» (Kool et al. 2013), con lo que llegamos a ver que la expectativa de recompensa es el basamento aquí: la más reciente revisión de estudios concluye que «la investigación en animales y humanos ha demostrado la contribución de los circuitos corticoestriatales y la dopamina a (...) la percepción del control» (Ly et al. 2019). Con esto se sugiere una tentativa neurobiología de la ilusión de control como el sistema de ciertos aspectos cognitivos y emocionales:
Junto con el rol central que se presume sobre la percepción de control en muchos aspectos de la vida humana, consistentemente ha sido encontrado que poseer un sentido de control tiene efectos adaptativos. Percibir control está asociado al bienestar emocional, a la reducción del impacto fisiológico de los estresores, la capacidad mejorada para lidiar con el estrés, la mejora del rendimiento, la disminución del dolor, y a una mayor probabilidad de hacer difíciles cambios conductuales (Thompson & Spacapan 1991). A través de una variedad de entornos desde el salón de clase al trabajo al centro médico, y en diversas poblaciones desde niños a adultos mayores, es generalmente adaptativo tener un sentido de control.» (Thompson & Schlehofer en Shah & Gardner 2008 p. 41)
- la «detección de errores» (córtex cingulado anterior —CCA: Bush et al. 2000),
- la ansiedad y el sistema límbico (hipocampo, amígdala),
- el automonitoreo y el control ejecutivo (córtex prefrontal),
- el aprendizaje, la memoria, y la expectativa de recompensa dopaminérgica: Wise 2004; cf. Robbins & Everitt 1996; Salamone & Correa 2012.
Entonces, la neuroanatomía, neurofisiología y psicología mencionadas subyacen a la consumación conductual enfocada a responder a tales impulsos motivacionales envueltos en el control, con sus respectivas características particulares. P.ej. las «zonas placenteras» o de expectativa dopaminérgica del cerebro subsirven tanto a la consecución de comida como de pareja, y están involucradas en la adicción a las drogas (Coria-Avila et al. 2008). La motivación del control, asociada causalmente al PM, tiene pues profundas raíces evolutivas y filogenéticas, consistentes a través de las especies animales en una compleja interacción endocrino-inmuno-neuro-conductual, con un impacto en el éxito reproductivo como más adelante veremos, p.ej. con la testosterona (T) motivando conductas de dominación y directamente asociada a la calidad del semen, o la oxitocina (OT) (Schultheiss & Brunstein 2010 p. 279) y la arginina vasopresina (AVP) (en animales: Robinson et al. 2008 p. 5; y en humanos: Walum et al. 2008) motivando la socialización y directamente el emparejamiento. Importantemente, los efectos de tales moléculas, entre los mencionados, resultan en la regulación del estrés, que impacta negativamente pues en el sistema reproductor y otros sistemas vitales como se mostrará también. La socialización y el estrés están enlazados: la dinámica de la interacción social se asocia causalmente al segundo (p.ej. la pérdida de dominación social es ansiogénica y estresora), del mismo modo que sus logros adaptativos lo regulan (p.ej. la inclusión social es ansiolítica y anti-estrés). Además, también de importancia para este artículo, estos efectos se dan bajo la interacción de aquellas con otro grupo de moléculas, las catecolaminas: adrenalina, noreprinefrina y dopamina (DA), donde la última, también involucrada en la socialización, parece regular corticoprefrontalmente el PM, como parte de una regulación (al menos prefrontalmente) de la agentividad, en tanto sabemos que la agencia es el fundamento de la cognición social (Frith & Frith 2010) y por tanto capital en la interacción social (Decety & Lamm 2007; cf. Bandura 2001).
Es decir que hay una relación entre socialización, estrés, y PM, con una base orgánica, que es ciertamente un sistema cerrado: el PM mismo como agentividad y causalidad es un producto de la cognición social traslapada con la visuoespacialidad (en palabras de Guthrie en Pyysiäinen & Anttonen 2002 p. 45, citando a Burkert: «tratar con el mundo en general como si fuera social y comunicativo», ver aquí); y probablemente a su vez esto resultó del crecimiento de los grupos sociales y de la complejidad socio-conductual que ello acarrea (p.ej. Allman et al. 2002 pp. 343-344: véase aquí). Dudo que haya evidencia directa de que el PM potencie la reproducción (como efecto de la socialización, lo que representaría su argumento evolutivo), sin embargo hay al menos una relación íntima entre el impacto del estrés en el sistema inmune y reproductor y su manejo incluyendo ‘efectos últimos’ corticales y de carácter regulatorio (de ‘arriba hacia abajo’) típicamente cognitivo-hiperagentivos (PM). Siendo así, se predice que el PM debería asociarse a un ciertamente específico correlato neurobiológico bajo algún grado de regulación genética. Al parecer así puede ser, siendo lo que se buscará sugerir en lo que sigue. Aún este grado de determinismo biológico (que no implica que el PM sea un aspecto absolutamente genético —p.ej. la evidencia de universalidad también indica variaciones culturales: Wilson & Keil 2001 p. 504—) está todavía por verse si en último término es estrictamente genético o epigenético, o, como más probable, ambos. P.ej. F.H. Previc es un caso particular de esta aproximación neurobiológica, mediante una teoría epigenética de la conducta y el intelecto humanos en general (Previc 2009, aquí se puede obtener el libro entero) que incluye la religión (Previc 2006), teniendo a la expansión del sistema dopaminérgico durante la hominización y los resultados de sus funciones como eje central de teoría (p.ej. las completas funciones de la DA y sus implicaciones evolutivas no las conocemos —hay evidencia que sugiere que sea mediadora de la neurogénesis cerebral animal: Berg et al. 2011).
Uno de los resultados de nuestras propias investigaciones en la bibliografía científica es que, ciertamente, una enorme cantidad de datos colocan a la DA (y las zonas frontales del cerebro donde es el más importante neuromodulador) como un importante factor fisiológico, sino el más importante, de la capacidad de abstracción en la cognición humana. Previc (2006) argumenta que la percepción del espacio que nos circunda y la interacción con él responde a ciertos sistemas percepto/cognitivo/conductuales dominados por la DA, siendo el «espacio extrapersonal», esto es el foco en el espacio y el tiempo distantes, el rasgo más notorio del intelecto humano, y el fundamento de la religión. Esto resuena fuertemente con el concepto MTT, al que se le ha dado importancia en cuanto a la religión (Suddendorf & Corballis 1997; cf. Bulbulia 2009 aquí o aquí) y a la cognición humana en general (cf. Suddendorf 1994; Boyer 2008; Suddendorf, Addis & Corballis 2009; Corballis 2019). De hecho, las bases neurales de esos sistemas (ibid. p. 504, Fig. 2), son consistentes con las del MTT (observación mía), compartidas en medida importante por la agencia y la Teoría de la Mente (véase Fig. 1 infra para una comparación). A diferencia de otros importantes neuromoduladores con los que ella misma interactúa (p.ej. serotonina —5-HT— o norepinefrina), la DA es ampliamente reconocida como el fundamento de la expectativa de recompensa, y consistentemente, está envuelta en la incertidumbre, probabilidad y predicción (Fiorillo et al. 2003; Aron et al. 2004, 2005; Niv et al. 2005; Schultz 2007; Glimcher 2011; Diederen & Fletcher 2020). En tanto juega este papel, crucial para la evaluación de estímulos, circunstancias y la toma de decisiones, la DA debería estar implicada prefrontalmente en la formación de las creencias, el autoconvencimiento y la convicción: efectivamente hay un rol relevante de la recompensa en las creencias generales (Harris et al. 2007) y específicamente religiosas (Harris et al. 2009, ver «CORRELATOS NEURALES DE LAS CREENCIAS RELIGIOSAS Y NO RELIGIOSAS»).

Fig. 1. Retomando la Fig. 2 de la Parte 1, compárese (A) la red perisilviana del hemisferio derecho, sugerida como especializada en orientación espacial y exploración —mientras su contraparte izquierda se especializa en el lenguaje (Karnath 2009; Gharabaghi et al. 2009), (B) la neurogenética de la red por defecto (Glahn et al. 2010), sobre lo que se ha hecho notar su traslapado con la agencia y el MTT, (C) la prospección (Buckner & Carroll 2007), (D) la ToM (ibid.), y (E) la agencia propiamente dicha entre otras funciones de autoprocesamiento (Blanke & Arzy 2005), con (F) el sistema extrapersonal, enfoque + acción (extraído de Previc 2006 p. 504, Fig. 2 aquí o aquí).
AVP y OT conforman una familia de péptidos muy similares molecular, genética y neuro-funcionalmente. «Comprenden circuitos neuroendocrinos que van desde ser conservados evolutivamente a evolutivamente distintos» entre mamíferos y aves, cuya modulación combinada en el cerebro y las estructuras neuroanatómicas que la soportan, integra «un factor determinante de la conducta de aproximación-evitación, y puede ser un objetivo prioritario de la selección natural relacionada con la sociabilidad» (Goodson 2008). «Los mecanismos neurales responsables de los efectos de la oxitocina y la vasopresina sobre la conducta social se considera que involucran algunos de los mismos circuitos (el núcleo accumbens y el pálido ventral) envueltos en el comportamiento relacionado con la recompensa. Estas regiones del cerebro son también componentes del sistema de recompensa de la dopamina» (Charney 2004 p. 208). En medio de la creciente aproximación bioquímica a la conducta, cabe decir que el hallazgo de esta neurobiología social no debería implicar una concepción reduccionista de ella, p.ej. considerando un modelo teórico en el que «los efectos de la oxitocina están restringidos por los rasgos de las situaciones y/o los individuos» (Bartz et al. 2011). Entre tanto, la AVP se ha descubierto como uno de los mediadores primarios y centrales de la respuesta de estrés (Chrousos & Gold 1992 p. 1246; Bartanusz et al. 1993; Tsigosa & Chrousos 2002 p. 866; Chrousos 2009; Beurel & Nemeroff 2014); está vinculada a la OT en los trastornos de ánimo (depresión: Purba et al. 1996), y contribuye junto a la DA y la 5-HT a la variación en la emotividad, el afrontamiento del estrés, las funciones cognitivas y las conductas sociales (Touma 2011).
OT funciona simultáneamente como fundamento del apego social y «un mecanismo biológico subyacente a los efectos estreso-protectivos de las interacciones sociales positivas» (Heinrichs et al. 2003). Esa función resulta ser bastante específica y directa neuroquímicamente: «la oxitocina parece emitir su señal a través de un solo receptor, una proteína diseñada para reconocer su forma y vibrar consecuentemente cuando se juntan, la dopamina y la serotonina, por el contrario, cada una tiene cinco o más receptores asignados a su cargo» (Angier 2009). Por supuesto, hay que decir que el apoyo social en general, es decir, la integración del individuo estresado a interacciones directas dentro de un grupo social funciona como potenciador del bienestar o como amortiguador («buffer») del estrés (Cohen & Wills 1985); mientras que aún la ilusión de apoyo social afecta positivamente el amortiguamiento del estrés (Vollmann et al. 2011); todo esto pues oxitoninérgicamente mediado (Heinrichs et al. 2003; Quirin et al. 2011; Simeon et al. 2011). En tanto que la OT incrementa las conductas de apego y afiliación social, p.ej. «comportamiento sexual, emparejamiento madre-lactante y adulto-adulto y memoria/reconocimiento social», se ha mostrado que tiene efectos positivos en el autismo, precisamente debido a que este desorden involucra trastornos de comunicación social y ensimismamiento (Bartz & Hollan 2008; Neumann 2008; Green & Hollander 2010). De hecho, se ha mostrado una relación entre el gen receptor de la OT (OXTR) y la cognición social, en la variación interindividual del perfil de empatía y la reactividad al estrés (Rodrigues et al. 2009), y como influencia en trastornos de la comunicación social (Park et al. 2010). Consistentemente con sus funciones de apego, tanto entre animales como entre humanos, la OT potencia la confianza, pero no como parte de una general toma de riesgos, sino que «específicamente afecta la disposición de una persona para aceptar riesgos sociales que surgen a través de las interacciones interpersonales» (Kosfeld et al. 2005). A esto se asocian p.ej. el mejoramiento del reconocimiento emocional (Guastella et al. 2010), regular la reacción de la amígdala a las caras emocionales (Domes et al. 2007a), e incrementar la generosidad (Zak et al. 2007). De hecho, la OT potencia la inferencia de estados mentales (Domes et al. 2007b); mientras que OT/DA/5-HT posibilitan una protección social contra el estrés (Tops et al. 2014).
Las monoaminas son neuroquímicos relacionados molecular y filogenéticamente que incluyen a las histaminas, las catecolaminas previamente mencionadas (adrenalina, noreprinefrina y DA), triptaminas (5-HT, melatonina) y otras monoaminas alcaloides (alucinógenos como psilocibina, DMT). No parece coincidencia que la DA y la 5-HT sean los más importantes neuromoduladores implicados en las ideaciones y experiencias mágico-místicas: su parentezco molecular con los alucinógenos propiamente dichos les hace tener no solo una natural relación sinérgica con ellos, sino que por sí mismos ejercen efectos «mágicos» ciertamente similares (p.ej. juegan papeles importantes en las alucinaciones psicóticas, que también son similares a los efectos de la psilocibina o el LSD: Vollenweider & Geyer 2001). Entre tanto, OT y DA tienen una demostrada interacción conductualmente relevante, y se han observado asociadas en el núcleo accumbens respecto al emparejamiento monógamo y la conducta maternal en animales (Liu & Wang 2003; Shahrokh et al. 2010), como también en humanos. «Así, los estudios a través de un rango de especies sugieren que las diferencias individuales en las conductas maternales derivan de la variación dentro de los sistemas mesolímbicos oxitocina-dopamina» (Shahrokh et al. 2010 p. 2284). Paralelamente, OT/DA están involucradas en la adicción a la droga, de hecho evidencia de ello en animales «apoya el argumento de que las vías neuronales y los mecanismos bioquímicos implicados en la adicción a la droga probablemente también median los refuerzos producidos por las recompensas naturales» (Liu & Wang 2003 p. 542); teniendo claro que la DA es ampliamente reconocida como mediadora de la expectativa de recompensa en humanos y demás primates (p.ej. Haber & Knutson 2009), la cual es un específico sistema neuroanatómico, fisiológico y psicológico del que es parte la OT (Ritsner 2010 p. 619; Charney 2004).
Es de enorme interés que el data acumulado en neurociencia social muestre que el sistema dopaminérgico de recompensa y la interacción social estén asociados, a nivel hormonal y genético (Caldú & Dreher 2007 aquí o aquí). La revisión de una amplia cantidad de estudios que realizan estos autores muestra que el sistema endocrino-gonadal femenino (estrógeno, progesterona), tanto entre animales y mujeres p.ej. durante el ciclo menstrual, afecta la activación dopaminérgica, incrementándose durante la fase folicular respecto a las regiones orbitofrontal, frontopolar, amigdalar y gangliobasal. Las reacciones emocionales también varían pre y posmenstrualmente, respecto al córtex orbitofrontal, amígdala y CCA; mientras que las preferencias por rostros de hombres con rasgos masculinos acentuados aumenta en la fase folicular, además de mayor atribución de posesión de recursos a hombres con fisionomía «dominante» (p.ej. mandíbula más cuadrada, menor distancia entre las pupilas y la frente). «Desde un punto de vista evolutivo, el aumento de actividad observada durante la fase folicular puede ser la base de la mayor disponibilidad, receptividad, y el deseo durante el período ovulatorio, que se piensa facilita la procreación» (ibid. pp. 57-59). Respecto a los hombres la vinculación entre recompensa, interacción social y T cuenta con menos estudios, y aunque ya hemos visto que la T se asocia a la dominación, hay que agregar que se vincula al procesamiento de estímulo sexual visual en hombres, con incremento funcional en el córtex orbitofrontal e insular, y al mejoramiento de las habilidades espaciales en mujeres (considerando que los hombres tienen relativa ventaja en ello). Similarmente a las hormonas femeninas, la T y el cortisol influyen en la toma de decisiones. Además este último afecta negativamente la confianza durante la interacción social bajo estrés, «sugiriendo que los sujetos con alta confianza interpersonal tienen menor activación del eje hipotalámico-pituitario-adrenal cuando expuestos al estrés social» (ibid. pp. 59-60). Este eje y su fisiología, que incluye al sistema límbico, es la base neurobiológica de la respuesta de estrés.
En la Parte 1 de este artículo se refirieron varios estudios mostrando que el genoma y la conducta humana están asociados. P.ej., la amígdala y el procesamiento emocional a través del reconocimiento facial y el seguimiento de la mirada, fundamentos de la cognición social (asociados al sistema de agencia), se relacionan a cierta región del cromosoma X (Skuse 2006). Entre tanto, hay fuerte evidencia conseguida mediante animales de la interacción entre genes y conducta social. Esta interacción tiene varias formas direccionales de influencia, como ‘socio→genética’: p.ej. activación de la expresión de genes precisos en el cerebro del pinzón ante el reconocimiento del canto de un congénere, expresión neurogenética en cuestión de minutos en el pez cíclico ante la circunstancia de ascenso jerárquico-reproductivo, o alteraciones neurogenómicas masivas en la abeja en su cambio de labor en la colmena (Robinson et al. 2008 pp. 2-4); ‘socio→epigenética’: transmisión generacional del estilo materno en ratas (descendencia menos estresada y más maternal que la que recibió menos cuidado maternal); y ‘eco→genotípica’, observada en las hormigas de fuego y aún en humanos, aunque no uniformemente, respecto a desórdenes vinculados a las conductas sociales como el autismo y la depresión (ibid. p. 5). Por supuesto, también hay una dirección ‘genético→social’: p.ej. entre dos diferentes especies de la mosca de la fruta se ha realizado la transferencia de un gen que controla el «canto» de apareamiento específico de cada especie, resultando en que una especie produzca el canto de la otra; mientras que comparando la expresión en el cerebro del gen V1AR receptor de vasopresina en dos especies de ratón de pradera, una monógama y otra polígama, se mostró que este gen controla las preferencias de emparejamiento; y los polimorfismos del gen FOR están involucrados en las diferencias inter-individuales en la mosca de la fruta respecto a la búsqueda de alimento, siendo que puede estar extendido entre los insectos sociales. En vertebrados incluyendo a los humanos, este gen y además el FOXP2 se han mostrado influyendo en la comunicación social, estando aparentemente implicados en la formación de los circuitos y las estructuras neurales subyacentes a diversas conductas sociales (ibid. pp. 5-6).
Partiendo de que la recompensa y la interacción social ambas ocupan regiones dopaminérgicas, una importante línea de investigación enfoca la base genética de la transmisión dopaminérgica en relación a las diferencias entre individuos y las patologías de ella, mostrando una correlación con ciertos rasgos de la personalidad y conductas vinculadas a la recompensa. Aquí están implicados los genes DRD2/DRD4 (receptores de DA), COMT (codificador de transmisión dopaminérgica) y DAT (transportador de DA). El DRD4 se asocia a la búsqueda de novedad y a la ludopatía, mientras el DRD2 a la drogadicción (ibid. p. 62). Como de esperarse, la ludopatía con su ilusión de control sobre el juego, y otras creencias irracionales, está asociada al PM (Lund 2010; Goodie & Fortune 2013). Así tenemos que «la oración, la meditación, el amor romántico inicial, y el abuso de drogas pueden tener en común la activación de las vías dopaminérgicas mesolímbicas del cerebro y la generación de intensos estados emocionales. En este sentido, la dependencia de un Poder Superior puede funcionar como un sustituto de la adicción» (Sussman et al. 2011). COMT, envuelto en la degradación de DA, y DAT, en su recaptación, interactúan regulando la anticipación de la recompensa y la activación del córtex prefrontal dorsolateral/ventral-orbital, amígdala, hipocampo y estriado ventral para ello. La expresión de ambos genes en las regiones gangliobasal y prefrontal regula los niveles de DA, cuyos muy altos o muy bajos niveles afectan las funciones ejecutivas y las respuestas emotivas ante la recompensa. Por ejemplo el aumento de DA durante el estrés reduce la función inhibitoria del córtex dorsolateral, pudiéndolo poner «fuera de línea» (Datta & Arnsten 2019); los mecanismos neuroquímicos que encontraron estos autores apuntan directamente a la regulación dopaminérgica del pensamiento mágico y la latencia de éste: en tanto se debilita la inhibición dorsolateral el PM domina la consciencia (recordemos la misma condición en el Alzheimer). Además este desbalance de la DA se vincula a las conductas repetitivas y compulsivas (de búsqueda o sobrestimación de recompensa) o a la esquizofrenia (Caldú & Dreher 2007 pp. 62-64).9 Un evidente nexo entre estos trastornos, p.ej. las adicciones con o sin sustancia (a las drogas, a los juegos, al internet), lo conforma la ilusión de control (Hyman 2007; Ladouceur & Sévigny 2005; Chak & Leung 2004), y la capacidad de expectación de recompensa puramente abstracta, asunto ya visto líneas atrás.
3.3. CIERTOS NEUROMODULADORES Y SU GENÉTICA: PENSAMIENTO MÁGICO Y EXPERIENCIAS EXTRAORDINARIAS
En este punto es necesario retomar la sección «Dopamina, pensamiento mágico, correlatos neurales de la creencia sobrenatural» del artículo «NEUROCIENCIA DE LA RELIGIÓN (II)...», para enfocar al gen COMT. Como allí mostrado, en principio la hiperdopaminergia está envuelta en los trastornos de la cognición espacial, en delirios y alucinaciones, en tanto que extensiones patológicas del PM: «las creencias sobrenaturales son un rasgo importante de los trastornos psicóticos de la manía y la esquizofrenia [...] incluso los adultos que no presentan desajustes mentales psicóticos en toda regla, los llamados casos “borderline”, se ha mostrado que mantienen un fuerte super-sentido.10 Estas creencias se llaman ideación mágica» (Hood 2009 pp. 234-235 —libro completo disponible aquí). P.ej., alteraciones en los ganglios basales, una fuente de DA e implicados en el control motriz, se asocian causalmente al Parkinson (Obeso et al. 2008); y su tratamiento, básicamente consistente en reactivar la DA, puede causar síntomas psicóticos incluyendo alucinaciones (Fénelon et al. 2000; Moser et al. 2003). Hay, adicionalmente, una consistencia en la lateralización hiperdopaminérgica hacia el hemisferio derecho entre PM, esquizotipia11 y esquizofrenia: «la ideación esquizotípica y la paranormal ambas se basan en una sobre-dependencia respecto al hemisferio derecho» (Pizzagalli et al. 2000). Incluso dicho desbalance hemisférico se postula como indicador del grado de PM, y viceversa, éste como indicador del normal desbalance hemisférico derecho de la DA. Hay en efecto específica y directa evidencia de que el PM (~ pensamiento paranormal: Brugger & Mohr 2008) está asociado a la DA (Krummenacher et al. 2002; 2010: un nivel elevado de DA «disminuye la sensibilidad en las decisiones percepto-cognitivas, pero sólo en los escépticos, y hace a los escépticos menos [escépticos] y a los creyentes [en creencias paranormales] un poco más conservadores» —esto detectando señales sensoriales en el ruido) y la regulación del gen COMT (Raz et al. 2008; quienes por cierto aclaran que que éste no debe ser tomado como «el gen» de la creencia paranormal).12 «Una mayor disponibilidad de DA, determinada por un número decreciente de haplotipos de alta actividad COMT, está asociada, en primer lugar, a una tendencia más fuerte hacia las creencias infundadas», mágicas y paranormales (Schmack 2015).
De hecho, el COMT, junto al gen T102C (serotoninérgico) con el que interactúa, han sido asociados al rasgo de personalidad de ‘absorción’, +una disposición a experimentar estados alterados de consciencia caracterizados por una atención intensivamente enfocada», que incluye vívidas imaginaciones y experiencias místicas (Ott et al. 2005). Consistentemente con esto, diversos estudios indican una interacción DA/5-HT asociada al PM y las alucinaciones y experiencias místicas y religiosas, que antes hemos tratado en «NEUROCIENCIA DE LA RELIGIÓN (IV): EL PAPEL DE LA SEROTONINA». Allí se mostraron algunos estudios sobre el impacto de los enteógenos en el cerebro, que sugieren que la 5-HT se asociaría más a las experiencias extraordinarias y las alucinaciones, con sus correlatos de fuerte fluctuación del estado de ánimo (uno de los atributos serotoninérgicos clásicos: Tiihonen et al. 1997; Heinz et al. 2001; como simultáneamente dopaminérgico: Burke et al. 2011), mientras DA al PM. De hecho, la 5-HT fue el primer componente fisiológico-conductual ligado a la religión (Mandel 1980 cf. Austin 1999 p. 288). Dado todo esto, la modularidad de la autotrascendencia evidenciada por ciertos estudios (Newberg et al. 2002; Johnstone & Glass 2008; Urgesi et al. 2010; Schuerbeek et al. 2010) se ha sugerido asociada a una probable base neurogenética (Urgesi et al. 2010). Los alucinógenos triptamínicos, a cuya familia química pertenece la 5-HT, interactúan con ésta como antagonistas (Titeler et al. 1988; Kim et al. 2011); se encuentran de modo natural en plantas y animales incluyendo al hombre y «han sido usados para propósitos religiosos y otros por centurias», comportándose más específicamente como ‘sustratos’, es decir son transportados por el transportador de 5-HT (SERT) y el transportador vesicular de monoamina (VMAT2) (Cozzi et al. 2009), que son proteínas reguladas genéticamente.
Que se encuentre uno o varios genes relacionados a la autotrascendencia es, en vista de los datos recientes de Urgesi et al. 2010, algo altamente probable y no deberá sorprender que así ocurra, lo problemático (además de las deficiencias implícitas que pueda tener un estudio) sea el modo en que los propios investigadores popularizan sus hallazgos, tal como pasó con el «gen de Dios» de Hamer. Ciertamente, el VMAT2 no es tal cosa, sin embargo este como otros genes vinculados a los neurotransmisores del grupo de las monoaminas (p.ej. DA, 5-HT) juegan de hecho un papel en la neurofisiología subyacente a la cognición religiosa. Aún así, este papel pudiera ser menos importante que en otras funciones orgánicas del todo distantes de la cognición religiosa (p.ej. el VMAT2 asociado a mastocitosis: Anlauf et al. 2006; o a la diabetes: Saisho et al. 2008). Por tanto, aunque es plausible la idea de que en las personas existan propensiones naturales hacia la autotrascendencia o espiritualidad, el papel de los genes bien parece ser de índole generalista y habría que buscar sus roles más o menos específicos en los subcomponentes cognitivos implicados. Aunque si bien es cierto no hay una división clara 5-HT/DA respecto al PM o a la religiosidad, es un hecho, por un lado, que tales neuromoduladores están envueltos en las experiencias extraordinarias (tanto místico-religiosas como místico-seculares), la autotrascendencia~espiritualidad y el PM (que sabemos ambos pueden ser independientes de la religiosidad), y por otro, que tales experiencias mismas juegan un papel modulador de la ansiedad y el estrés.
El estudio del gen COMT abre la puerta a un camino intrigante de hipotetización sobre la cognición humana en general (p.ej. a partir de Goldman et al. 2009; o como explícitamente lo hace Previc 2009), y el pensamiento mágico en particular. A saber, este gen es pues un candidato para la esquizofrenia, e interactúa con otros genes candidatos, en razón de sus funciones (degradación de DA) como regulador de la eficiencia del córtex prefrontal (Nicodemus et al. 2007). Adicionalmente, la evidencia acumulada apunta a un traslapado genético esquizofrenia/autismo (Van Winkel et al. 2010; Gejman et al. 2010).
3.4. LA INTERACCIÓN SOCIAL: ESTRESORA, A LA VEZ QUE REGULADORA DEL ESTRÉS
Como especie social, o más apropiadamente, hipersocial (Boyer 2000), o aún ultrasocial (Haidt 2011), el ser humano debe ser hipersensible al estrés en todo lo que respecte a falta de interacción, competencia y dominación social, y no es sino recientemente que podemos asegurar científicamente que existe incluso una experiencia de «dolor físico» ante algo como el rechazo social (Eisenberger et al. 2006; involucrando el córtex cingulado anterior, insular y somatosensorial: Kross et al. 2011; ver también Yao et al. 2019). El rechazo social, literalmente, duele («distress») y, consistentemente, hay evidencia empírica previa p.ej. del rechazo experimentado por los gays como estresor cual ilustración específica (Meyer 1995; 2003); mientras que genéricamente el rechazo social es un estresor, con la respuesta sujeta a dimorfismo sexual hacia una hiper-reactividad masculina (Kudielka & Kirschbaum 2005 abstracto / completo). Implicando rechazo (y dolor «distress») social y/o factores adicionales, eventos como las discusiones, el divorcio, el nacimiento del primer hijo, el embotellamiento de tráfico, el desempleo, la eficacia laboral, una visita inesperada o la destrucción del hogar por un desastre natural, son estresores (Thoits 1995 p. 54; Willis et al. 2018). Y, si por un lado tenemos que el PM es identificable en aspectos sociales, políticos o económicos, de acuerdo con la relación PM/estrés, por otro tenemos que precisamente aspectos de organización social también son estresores (p.ej. Aneshensel 1992). No sería necesario hacer una búsqueda exahustiva en la literatura psicológica/sociológica (para ello ya se han referido estudios pertinentes) sino comprobar alrededor nuestro que buscar superar o evadir tales situaciones, está frecuentemente asociado al fetichismo, la superstición o a la religión (digamos: «¡deséame suerte hoy!», «Dios sabe porqué pasa esto»).
Entre tanto, «los dos recursos personales de afrontamiento [del estrés] más frecuentemente estudiados por los sociólogos son un sentido de control o dominio sobre la vida (p.ej. un lugar interno o externo de orientación del control) y, menos comúnmente, la autoestima» (Thoits 1995 p. 60); mientras que el apoyo social es a la vez moderador y mediador del estrés (Erin et al. 2014; Costa et al. 2017; Viseu et al. 2018; Polizzi et al. 2020). Con todo esto el ser humano debería poseer entonces hiperactivos dispositivos anti-estrés, que involucren mecanismos neurocognitivos no vistos en otras especies, y esto necesariamente, en tanto que la naturaleza híper o ultrasocial humana es consecuencia de su encefalización. Efectivamente, «ser miembro de un grupo muy unido tiene directos beneficios para la salud mental, aunque esto no actúa como un moderador sobre los resultados del estrés; la percepción de apoyo emocional (sea real o no) tiene efectos directos y amortiguantes sobre la severidad e importancia de los eventos vitales negativos para los resultados de estrés»; y finalmente, «tener una relación que anima a la confianza íntima en otros tiene el mayor efecto atenuante sobre los resultados del estrés» (Lackey 2006)*. De acuerdo con el referido Haidt (2011), de las no muchas especies ultrasociales (p.ej. abejas, hormigas, termitas), en todas el mecanismo es la consanguinidad absoluta (todos los individuos tienen afinidad fraternal y son hijos de una sola hembra), sin embargo, solo una especie también ultrasocial, el hombre, muestra otro mecanismo de apego social, que es único y que no es de parentesco fraterno-genético, sino imaginario y abstracto:
«[...] nuestra habilidad de forjar equipo alrededor de objetos y principios sagrados. [...] Gente de toda fe está unida por su devoción compartida a objetos, personas y principios que se consideran sagrados. Esta habilidad es crucial en la guerra. Y en la política. Somos igualmente buenos para unirnos en equipos, especialmente al competir contra otros».
Mientras que en «LO QUE NOS DISTINGUE DE LOS SIMIOS», se mostró que la ToM es un proceso fundamentalmente abstractivo, o en todo caso, alcanza espontáneamente elevados niveles de abstracción. Ahora, si bien es ampliamente reconocido el papel que juega la DA a través del córtex prefrontal, como vimos también respecto a sus funciones ejecutivas, cognición social y procesamiento emocional, no hay datos explícitos de la vinculación ToM-DA, aunque es bastante obvia en varias líneas de investigación como las consultadas aquí o en el artículo que se acaba de referir. En todo caso, hay algunos estudios sugerentes sobre el desarrollo de la ToM que enfocan ciertos trastornos asociados a ello, en los que está involucrada la DA (p.ej. Carlson & Moses 2001; Sabbagh et al. 2006). Y, teniendo hasta aquí una robusta asociación neuropsicológica entre PM, motivación de control y estrés, por último se enfocarán otros aspectos neuroanatómicos y fisiológicos de esta relación misma, necesarios de revisar, que conforman el así llamado sistema del estrés (que libera la respuesta de estrés).
3.5. NEUROBIOLOGÍA DEL ESTRÉS
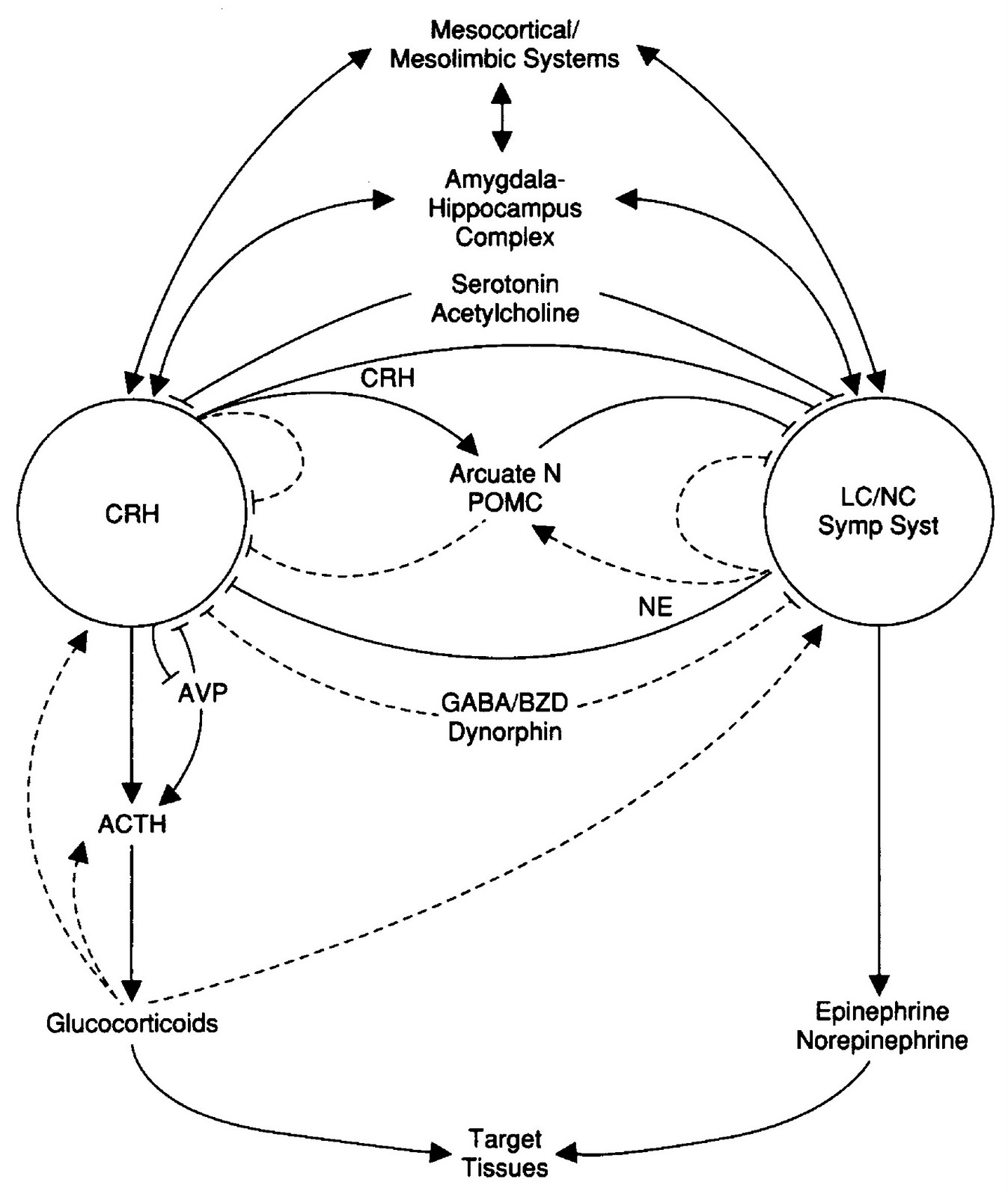
Fig. 2. Una representación simplificada y heurística de los componentes centrales y periféricos del sistema de estrés, sus interrelaciones funcionales, y sus relaciones con otros sistemas del sistema nervioso central envuelto en la respuesta de estrés. CRH: hormona liberadora de corticotropina, POMC: proopiomelanocortina, LC/NE Symp Syst: sistema locus ceruleus-norepinefrina/simpatético, AVP: arginina vasopresina, GABA: ácido gamma-aminobutírico, BZD: benzodiacepina, ACTH: corticotropina. (Extraído de Chrousos & Gold 1992 p. 1247 Fig. 1 —abstracto / artículo completo—)
En segundo lugar hay también, como dicho arriba, una neurofisiología específica que regula estas regiones. Dos principales sistemas neuro-endocrinos, corticotropina (hipotálamo) y norepinefrina o noradrenalina (médula), estimulados por la AVP, activan otros tres mayores sistemas:
- el dopaminérgico (mesocortical y mesolímbico) mediante la norepinefrina, que inerva el córtex prefrontal (especialmente el CPFM) y soporta los efectos motivación/refuerzo/recompensa (hay evidencia directa del envolvimiento de la dopamina en la respuesta de estrés: Cabib & Puglisi-Allegra 2012; Zhang et al. 2019);
- el amígdalo-hipocampal, también vía noradrenérgica, para procesar la información emocional del estresor y la activación de patrones fijos (memorizados) como el miedo condicionado;
- y la automodulación hipotalámica vía proopiomelanocortina, de los sistemas corticotropinérgico y noradrenérgico, ya en la fase final de la respuesta de estrés, activando los contraefectos analgésicos, de adormecimiento (activación endorfinérgica) y regulación emocional (Chrousos & Gold 1992 p. 1246; Tsigosa & Chrousos 2002).

Fig. 3. Representación simplificada de las interacciones entre el LHPA y otros sistemas vitales, entre ellos el eje reproductivo (A), el de crecimiento y tiroideo (B), y el sistema inmune (C). LHRH: hormona liberadora de hormona luteinizante, CRH: hormona liberadora de corticotropina, LH: hormona luteinizante, FSH: hormona estimulante del folículo, ACTH: corticotropina, GHRH: hormona liberadora de la hormona de crecimiento, STS: somatostatina, TRH: hormona liberadora de tirotropina, GH: hormona de crecimiento, TSH: hormona estimulante de la tiroides, T4: tiroxina, SmC: somatomedina C, T3: triyodotironina, IL-1: interleuquina 1, IL-6: interleucina 6, TNF: factor de necrosis tumoral, y PAF: factor activador de plaquetas. (Extraído de Chrousos & Gold 1992 p. 1248 Fig. 2).
Por un lado, lo importante de este hallazgo es que sugiere una asociación neurológica entre el estrés y el PM más allá del CPFM, respecto a lo que llamo ‘eje posterior de agentividad’: el sistema conformado por las cortezas temporal/parietal, especialmente la JTP (abarcando más exactamente BA39/BA40/BA22: Christopher et al. 2010 p. 508), que es el sustrato neural, conjuntamente con el CPFM, de la agencia y la ToM (Frith & Frith 2010 p. 165: ambos construidos a partir del reconocimiento de movimiento biológico), el MTT (Arzy et al. 2008: «el MTT implica no solamente los mecanismos de la memoria, sino también los mecanismos relacionados a las imágenes mentales y la autolocalización») y la red por defecto (Christoff et al. 2009; ver arriba las imágenes comparadas de la Fig. 1). Aunque es necesario encontrar evidencia neurológica directa de que las personas, estando simultáneamente bajo estrés y manifestando PM, tienen mayor actividad temporo-parietal, la comparación de la evidencia de atribución de agencia a objetos y eventos inanimados, MTT y ya explícitamente creencias religiosas y experiencias místicas (todos aspectos del PM), conforma una fuerte sugerencia de esa asociación neurológicamente fundada estrés/PM. Por otro lado, este mismo eje tiene un funcionamiento multimodal que es neuralmente hiperactivo a muy temprana edad (desde inicios del primer año de vida), y por lo visto debido a ello es capaz de sostener, aún independientemente del CPFM (que de hecho tarda más tiempo en madurar), procesos de categorización perceptual implícitamente abstractivos (véase la sección «Agencia abstracta sin ToM, y alcance de la ToM hasta la agencia sobrenatural–religiosa» del artículo sobre neurodesarrollo, abstracción y ToM). Este sistema agencia+ToM+MTT, sugerido como el sustrato de la agencia sobrenatural, conformaría en sí mismo la base neurocognitiva del PM. Esto implica inmediatamente que PM y agencia sobrenatural no son sino lo mismo, lo que es coherente con la necesaria diferencia entre el PM y el pensamiento científico, estipulada arriba en la sección 1.2.

Fig. 4. Otra ilustración de las estructuras cerebrales, neurotransmisores, neuropéptidos y hormonas implicadas en el estrés. «Es evidente que el estado funcional de las regiones del cerebro como la amígdala (neuropéptido Y, galanina, hormona liberadora de corticotropina [CRH], cortisol y norepinefrina), el hipocampo (cortisol y norepinefrina), locus coeruleus (neuropéptido Y, galanina, y CRH), y el córtex prefrontal (dopamina, noradrenalina, galanina y cortisol) dependerá del equilibrio entre los múltiples inputs neuroquímicos inhibidores y excitadores». (Extraído de Charney 2004 p. 203).

Fig. 5. «La figura muestra un resumen simplificado de algunas de las estructuras del cerebro y la neuroquímica relevante de la mediación de los mecanismos neuronales de recompensa (trayecto púrpura), el condicionamiento y la extinción del miedo (amarillo), y el comportamiento social (azul) [...] se puede ver que hay una considerable superposición en las estructuras del cerebro asociadas con estos mecanismos neuronales. Esto sugiere que puede haber interacciones funcionales clínicamente relevantes entre los circuitos. Por ejemplo, un circuito de recompensa que funcione correctamente puede ser necesario para el reforzamiento de las conductas sociales positivas. Un circuito del miedo muy sensible o un alterado proceso de extinción puede influir negativamente en el funcionamiento del sistema de recompensa. [...] Alteraciones en un neurotransmisor, neuropéptido, o sistema hormonal afectará a más de un circuito. [...] El estado funcional de estos circuitos tiene una influencia importante sobre el estrés relacionado con la psicopatología y el descubrimiento de nuevas terapias». (Extraído de Charney 2004 p. 209).
En consonancia con todo esto, el CPFM (ventromedial) y el CPFDL juegan papeles de fundamental importancia aquí: como sustrato de las emociones (Krueger et al. 2009; p.ej. el grosor del córtex prefrontal dorsomedial se relaciona con «la tendencia a suprimir la expresión de las emociones»: Kühn et al. 2011), y de la formación y evaluación de creencias (Harris et al. 2007; 2009; p.ej. CCA y CPFDL se activan al detectar sesgos en las respuestas intuitivas: De Neys et al. 2008). Arriba vimos que la asociación Alzheimer/estrés/PM implica un desborde cognitivo-mágico (parecido a lo que ocurre con el Parkinson y su desborde de alucinaciones bajo tratamiento dopaminérgico), predeciblemente debido a alguna deficiencia en el papel inhibitorio del córtex prefrontal, y de hecho eso ha sido recientemente observado: una hipofunción en el CCA-derecho (recordemos: detección de errores), BA10, y también en la JTP-derecha, giro temporal inferior izquierdo, córtex somatosensorial primario derecho, estriado y cerebelo, correlativa a la falta de consciencia en pacientes con Alzheimer de sus propios déficits cognitivos (Amanzio et al. 2011). Esta hipofunción toma lugar en parte de las bases de la agencia, ToM, y MTT, mayormente lateralizado hacia el hemisferio derecho, mientras que el estriado es un núcleo dopaminérgico gangliobasal implicado en el control ejecutivo, lo que da cuenta de la neurobiología de la desinhibición ideacional y conductual (pero no sobre la pérdida de la memoria, lo más generalmente relacionado al Alzheimer —previamente se han identificado regiones más específicamente asociadas a la memoria en esta patología, como el hipocampo: Diagnostic Imaging 2009).
Estos resultados son interesantes porque ponen en relieve la necesidad de estudiar exclusivamente al PM en el cerebro, para, en un cometido explicativo profundo como el aquí pretendido, no depender de datos indirectos. P.ej., el Alzheimer nos está mostrando que el desborde del PM puede asociarse a una hipofunción predominantemente derecha (aunque estos estudios recientes deberán confirmase, ello es consistente con el concepto del hemisferio izquierdo como un «intérprete» elucubrador de explicaciones —obviamente compensando la hipofuncionalidad derecha), mientras se ha propuesto que el PM sea precisamente una consecuencia de la hiperactividad derecha (el «intérprete» del hemisferio izquierdo simplemente se vería abrumado), lo que conduce a la especulación de que el PM podría co-depender tanto de una promiscuidad mágica del hemisferio derecho, de una desaforada interpretación narrativa del izquierdo (que también desemboca en la elaboración de creencias irracionales), como del balance entre ambos estilos cognitivos. Ciertamente, la DA está implicada en todo ello y también es necesario estudiar cómo ella regula este balance, aunque ya hay algunos estudios evidenciando al PM como un asunto asociado más al hemisferio derecho, y aún a cierta base genética. En todo caso, la recompensa sería un importante factor común, implicado pues en las ideas y conductas repetitivas, más o menos independientes de los procesos inhibitorios, como al menos ampliamente mostrado mediante los efectos de las drogas, asunto que se puede identificar tanto en el ritual religioso (Alcorta & Sosis 2005) como en la obsesión sea culturalmente aceptada o patológica (Boyer & Lienard 2006; 2008). Prácticamente todo esto, además, esta asociado a la ansiedad y el estrés, y, siendo que esta integración CPFM-CPFDL implica la convergencia emoción/cognición/funciones ejecutivas, se ha sugerido anteriormente (ver aquí) que este proceso significa la integración del PM (ToM/MTT), la toma de decisiones y la ejecución motora: es decir, de aquí surge la conducta ritual enfocada en las creencias y agentes sobrenaturales, y por extensión, la religiosidad.
4. Discusión
Esta segunda parte sugiere que ciertas respuestas mentales y conductuales a los estresores no son una mera adquisición cultural, sino más bien manifestaciones de un estilo cognitivo biológicamente fundado pero modelado cultural/contextualmente. Dicho estilo es el PM, que justificadamente podemos caracterizarlo como intuición o cognición por defecto, lo que puede distinguirse, entre tanto, de las ‘creencias mágicas’ y la religión. En años recientes este punto de vista naturalista ha adquirido fuerza en la literatura psicológica (p.ej. Shermer, Hood, Subbotsky, Rosengren, Rozin, Markle, etc.). Siguiendo a Subbotsky aquí se asume que el PM, como «mente-sobre-materia» incluyendo «mente-sobre-mente», es ciertamente equivalente a ‘agentividad’, ‘mentalismo’, ‘mente inmaterial’, ‘espacio externo como agente’ (concepto propuesto en algunos artículos nuestros), ‘mental time travel’, ‘mente extendida’, y aún ‘espiritualidad’ (es decir ‘autotrascendencia’). No podría dejar de observarse que, por un lado, esto recuerda mucho al ‘animismo’, sobretodo cuando Guthrie (2002) lo define: «el sentido de atribución de agencia a objetos que no la tienen»; y por otro lado, que todos los conceptos mencionados pueden identificarse con el concepto de ‘sobrenatural’. Por antonomasia pues, todo esto es anti-científico, aunque no de modo absoluto o no en todos los casos en que se manifiestan los procesos cognitivos que los estructuran (p.ej. la mecanocuántica implica fundamentos auténticamente contraintuitivos que sin embargo son hechos empírico-científicos, y deben asimilarse; o también en los casos de creencias antiguas tenidas como religiosas o mágicas pero que ahora son realidades tecnológicas —como la comunicación a distancia). Mientras tanto, nada de ello implica que esos conceptos sean necesariamente equivalentes entre sí, aunque creemos que todos involucran la definición de Subbotsky. Por supuesto, es ampliamente aceptado que el PM es parte implícita e incluso un importante indicador de la esquizotipia y, ya cruzando el umbral del trastorno patológico de todos esos conceptos, de la psicosis.
Todos los conceptos mencionados, a su vez, juegan un importante papel tras la religión, pero en absoluto se trata de equivalentes a ésta. Por lo tanto, no es la religión lo que puede tener una base genética y ser el resultado de la selección natural, sino los mecanismos neurocognitivos referidos por esos conceptos (agentividad ~ mentalismo ~ mental time travel). Las correlaciones beneficiosas para la salud entre éstos y la religión o la religiosidad, no son suficiente argumento para extrapolar su probable origen adaptativo a las segundas. Además se tiene bien claro que las creencias mágicas y las supersticiones pueden ser potencialmente peligrosas, pero esto es algo que se espera en los operantes adaptativos en tanto su origen evolutivo estuvo restringido a cierto contexto u objetivo, y que no necesariamente serán beneficiosos todas las veces ni en todas las situaciones nuevas. Veamos por ejemplo evidencia de que «un sentido de propósito en la vida» y la espiritualidad (autotrascendencia) tienen fuerte correlación con el afrontamiento del estrés, más aún que el optimismo y el apoyo social (en un muestreo de individuos afroamericanos en EE.UU.: Doucette et al. 2008); lo que es consistente con otros hallazgos de correlaciones entre religión, espiritualidad y salud mental (ibid. p. 4; cf. Southwick et al. 2005). Esto sin dejar de considerar que del mismo modo en que la socialización es ambivalentemente estresora y anti-estrés, las creencias religiosas son también una fuente ansiogénica y estresora: «[la religión] [...] genera tanta ansiedad como la alivia: fantasmas vengativos, espíritus ofensivos y dioses agresivos son tan comunes como las deidades protectoras» (Boyer 2003 p. 120 Table 1). Esta ambivalencia es pues dramáticamente patente en las creencias religiosas, especialmente en las más doctrinales (recordemos los datos sociológicos que la correlacionan con la decadencia social), lo que apunta a que en realidad no hay genes de la religiosidad, sino que ella «depende de manera crucial de la cultura» (Banerjee & Bloom 2013). La religión, en un sentido importante, es un derivado histórico de la convergencia de la interacción social con el PM, lo que la capacita para manejar las interrogantes existenciales, los intereses sociales (que ella misma origina), y el estrés. El PM luce más como un dispositivo anti-estrés per se.
En tanto que estos aspectos (religión/espiritualidad) se fundan en el PM (el sentido de propósito es específicamente una forma de causación mágica), el PM mismo está vinculado al directo amortiguamiento del estrés, siendo específicamente esto lo que otorgaría el efecto anti-estrés a la interacción social y a la identidad grupal, como al menos ocasionalmente visto en la religiosidad y la religión (y muy probablemente también en la política, el arte, la economía de mercado o aún la medicina cuando son percibidos como asuntos trascendentales y/o cuasi-agentes causales). La sugerencia hecha en este artículo es robusta porque se apoya importantemente en que, como vimos sobre la recompensa (DA y regiones gangliobasales-prefrontales asociadas) y la conducta social (OT, AVP, DA, 5-HT, genes y regiones asociadas subcorticales, límbicas y prefrontales), y luego sobre DA/5-HT y las experiencias espirituales/extraordinarias o ya directamente el PM (sistema de agencia), es evidente que existen aspectos neurofisiológicos y neuroanatómicos regulados genéticamente que lucen intrínsecos al PM. Esto a pesar de que no haya una demostración explícita y directa de ello y aún dentro del incipiente estado en que se hayan estos conocimientos. Como dicen Raz et al., no hay un gen del PM. Entre tanto, el hecho último de que esos mismos aspectos biológicos formen parte de la respuesta de estrés es un fuerte indicio de una asociación PM-estrés, que ciertamente no ha sido aún abordada neurobiológica sino psicológicamente. Entre tanto, los notorios estudios de Brugger, Raz y Krummenacher sobre PM y DA, sí apuntan a confirmar un soporte neurobiológico común, asociado causalmente al PM por un lado, y a la respuesta de estrés por otro.
Más aún, si como parece el PM es agencia + ToM + MTT, esto es consistente con que el estrés también involucre la actividad inferoparietal y supratemporal, lo que a su vez conforma otra compleja línea de apoyo a la asociación PM-estrés. Como ya señalado, estas regiones, que además conforman la TPJ, estructuran un núcleo primario de agencia (p.ej. movimiento biológico y ToM), de procesamiento implícitamente abstractivo y sustrato fundamental de prácticamente todos los aspectos agentivos y ‘mágicos’ (es decir, extraordinarios: la JTP es el sustrato de la «duplicación ilusoria, autolocalización ilusoria, punto de vista ilusorio, y agencia ilusoria» —Blanke & Arzy 2005) asociados a las ideaciones y experiencias de PM y cognición religiosa. Boyer (2003, p. 122 Table 2) ha elaborado «un marco para una neurociencia cognitiva de la religión», fundamentada en una red cerebral temporo-parieto-frontal implícita en la agencia, el procesamiento de las señales sociales y la empatía. La existencia de aspectos neurocognitivos que sugieren una asociación, en sentido preparatorio y regulatorio, del PM con el estrés, además de consistente con el vínculo conductual arriba mostrado, refuerza este marco. Esto adicionalmente se respalda en que la JTP, y más ampliamente aquel sugerido eje temporo-parietal agentivo, por ser una región multimodal (procesa información visual, auditiva, somatosensorial, social y lingüística), al sufrir alteraciones funcionales o estructurales (p.ej. esquizofrenia) produce una gama de alteraciones perceptuales e ideacionales mediadas, tal como estipulado por Blanke & Arzy (2005), por la distorsión hiperagentiva, implicando consecuentemente trastornos en el lenguaje y delirios de control por fuerzas o entidades externas, es decir, una «anormal activación del sistema de percepción y comunicación social» (Wible et al. 2009). Dada la fuerte evidencia de asociación del PM, la esquizotipia y el psicoticismo, que nos muestra un continuum entre la autotrascendencia (espiritualidad), las creencias paranormales y los delirios esquizofrénicos, es de esperarse que los fundamentos cognitivos (agentividad) y neurales (sistema temporal/parietal—prefrontal) esten importantemente correlacionados, y en efecto esto es lo observado.
Luego se recordaron aspectos conectivos y funcionales del CPFM para observar que es ciertamente independiente del sistema temporo-parietal, o al menos no están directamente conectados. En consecuencia, debido además a la lenta maduración del CPFM (el BA10 —también incluido en el CPFDL), nos volvemos a encontrar con que el ‘eje posterior de agentividad’ sea pues un sistema multimodal categorial y abstractivo autofuncional, de cuyos procesos no somos conscientes, que con el desarrollo del cerebro posterior, a los 3 años de edad, se ve complementado por la funcionalidad integrada CPFM-CPFDL (BA10). Todo esto apunta a que las bases neurales del PM son prácticamente las mismas que las de la ToM, y por tanto también traslapadas con las del MTT. Por lo menos, en el ámbito académico se ha sugerido «una relativa hiperdopaminergia de el hemisferio derecho como la base biológica de la ideación mágica» (Mohr et al. 2003; ver también Narmashiri et al. 2019), mientras que su familiaridad con la psicosis involucra al córtex parietal (Mohr et al. 2003 p. 172) y la disfunción del sistema somatosensorial (Lenzenweger 2000). Recuérdese que en la Parte 1, como ilustrado nuevamente aquí en la Fig. 1, se mostró que agencia, ToM y MTT están sentadas en un sistema neuroanatómico bajo importante regulación genética (ver aquí), tanto en cuanto a su contectividad (materia blanca), como al córtex (materia gris). Y ahora se ha mostrado que el sistema dopaminérgico está regulado genéticamente. Interesantemente (ver aquí), la propia conexión entre el córtex frontal izquierdo y derecho, estando bajo mayor influencia ambiental que genética, explicaría la existencia de creyentes mágicos (relativa predominancia hemisférica derecha) y escépticos (relativa predominancia hemisférica izquierda) en tanto parecen diferenciarse meramente en el aprovechamiento del pensamiento analítico (Gray & Gallo 2016), un proceso lateralizado en el hemisferio izquierdo (lo que no significa que se reduzca a él), mientras que, en contraste, la conexión interhemisférica temporo-parietal está bajo mayor influencia genética, lo que daría cuenta de la persistencia inconsciente del PM (o de porqué la condición de estrés lo hace aflorar)
5. Conclusión
NOTAS:
1. Un ejemplo de este diálogo lo tenemos precisamente al buscar atender la pregunta «¿por qué los individuos pueden responder a la inseguridad personal mediante el incremento de su religiosidad?» (Rees 2009 p. 13): hay que tomar en cuenta pues los estudios del campo psicológico mostrando la asociación estrés ←→ falta de control ←→ pensamiento mágico ←→ religión. Entre tanto, véase en Epiphenom (el blog de este autor) otros indicadores socioculturales correlacionando inversamente con el grado de R, como mayor paz, menos corrupción, mayor desarrollo científico o menos desigualdad.
2. Véase a Shermer citando a Hood o esta entrevista a Hood (cf. Hood 2009 —libro completo disponible aquí). También véase PM identificado en análisis estadísticos: Elkan 2001; sobre la implementación de la informática en el campo de la salud: Diamond & Shirky 2008; en educadores: Genovese 2005; o en el tratamiento del autismo: Psychology Today 2009. Ver un análisis cognitivo del ateísmo en La descreencia en Dios es analítica ¿o no?.
3. En vista de que mi argumento consiste en que sencillamente, agencia, ToM y MTT por lo mismo que implican atribuir mentalidad a objetos inanimados y aún a eventos accidentales que evidentemente no tienen cerebros y por tanto no pueden tener mente, pueden así justificadamente categorizarse como un mentalismo sobrenaturalista, es interesante que Subbotsky (2010 pp. 142-143 —libro completo disponible aquí) plantee una dicotomía conceptual, pero patente y efectiva psicológicamente, entre la realidad física y la realidad social. A saber, a pesar de un persistente debate mente/cerebro, «esto no significa que la mente pueda existir sin el cerebro», ya que se trata de una verdad científica y, sobretodo, se trata de una verdad naturalista (exactamente lo mismo que observa Bloom 2004). Respecto al MTT, algo muy similar: es una verdad científico-naturalista la irreversibilidad del tiempo, sin embargo, podemos viajar imaginariamente tanto hacia atrás como hacia adelante en el tiempo (Tulving 2002 p. 2), violando, siempre en sentido conceptual (y/o psicológico), las leyes naturales. En ese mismo sentido, está claro que la agentividad viola la ciencia y el naturalismo: imputamos mentalidad a las montañas y a las computadoras para actuar adaptativamente de acuerdo a ello, es decir, esto forma parte esencial de nuestra naturaleza como primate hipersocial que somos. Ahora bien, si se trata de que científico-naturalistamente asumimos que lo sobrenatural no es más que un término vacío sin referente real, con un significado pura y exclusivamente gramático-conceptual, cual es precisamente mi postura existencial (aunque sabemos que la dimensión socioemocional del término, esto es, la convicción en ello —de que sí existe realmente— alivia la falta de control y el estrés), hace que la agentividad asbtracta caiga de lleno en tal concepto.
4. Traducción modificada del término en inglés «agenticity» —literalmente: «agenticidad»— de Shermer 2009: «la tendencia a creer que el mundo es controlado por agentes intencionales invisibles».
5. El concepto ‘dispositivo hiperactivo [o hipersensible] de detección de agencia’ —en inglés ‘HADD’, anacronismo de ‘Hyperactive Agency Detection Device’— fue introducido por J.L. Barrett: Barrett 2000.
6. «Look Who's Irrational Now» (Hemingway 2008, The Wall Street Journal). La nota periodística, de un aire irónico sobre la campaña neo-ateísta antirreligiosa, comenta sobre otros datos intrigantes: superstición y creencias paranormales incrementándose con el grado de estudios, y aún «autoproclamados ateos» creyendo en «una fuerza impersonal», rezando o creyendo en el cielo. Sin necesidad de proclamar estas estadísticas (posiblemente cuestionables, cf. Kosmin 2010) como alguna demostración de la prevalencia y carácter subyacente del PM, de todos modos son consistentes con la amplia evidencia psicológica.
7. Spelke & Kinzler (2007): «la cognición humana está fundada, en parte, sobre cuatro sistemas para representar objetos, acciones, número y espacio. Puede estar basada, también, en un quinto sistema que representa congéneres sociales. Cada sistema tiene profundas raíces en la filogenia y ontogenia humanas, y guía y da forma a la vida mental de los adultos. Convergente investigación con infantes humanos, primates no humanos, niños y adultos en diversas culturas puede ayudar a entender estos sistemas y los intentos de superar sus límites».
8. De entre los varios investigadores que desde los años 80s estudian la ToM, A.M. Leslie elaboró una hipótesis de estructuración jerárquica del Sistema de Agencia (Leslie en Hirschfeld & Gelman 1994 pp. 119-148; Leslie 1995). Aclarando que agencia no es sinónimo de animación ni de entidad biológica, sino el conjunto de propiedades que ciertos objetos poseen mientras que otros no, propone los subsistemas: «propiedades mecánicas» o «ToBy» —Teoría del Cuerpo— («los Agentes tienen propiedades mecánicas que los meros objetos físicos no tienen»), «propiedades de la acción» o «ToMM sistema1» —Teoría del Mecanismo Mental— («los Agentes actúan persiguiendo metas y reaccionan al entorno como resultado de su percepción» [...] «“causalidad teleológica”»), y «propiedades cognitivas» o «ToMM sistema2» («la conducta de los Agentes está determinada por propiedades cognitivas, p.ej. poseer cierta actitud hacia la verdad de una proposición» [...] «“causalidad psicológica”»). «Leslie argumenta que alrededor de los 6 y 8 meses los infantes alcanzan el sistema1, y empiezan a entender el rol que los objetos juegan en las acciones dirigidas a metas [...] en la etapa del sistema2, emergiendo entre 18 y 24 meses, los niños desarrollan la capacidad de pretender y entender las pretensiones de otros» (Cartwright 2008 p. 31). Entre tanto, como este autor señala respecto a Leslie, eventualmente se ha acumulado evidencia que no apoya plenamente una temprana habilidad de ToM (además los datos neurocientíficos sugieren que el núcleo matriz es la predisposición perceptual hacia el movimiento biológico y no precisamente hacia la física-mecánica-clásica, o en todo caso, estamos ante un único módulo «poli-nuclear» físico/biológico —y psicológico). Aunque tampoco podemos descartar a Leslie: p.ej., el amplio lapso de aparición de la ToM se extiende desde los 2.5 hasta los 5 años de edad, bajo el paradigma de la falsa creencia, mientras que alguna explicación debe haber de la evidencia sugiriendo, por lo menos, algún grado de agentividad pre-ToM (véase aquí, aquí, cf. Senju et al. 2011).9. Véanse más funciones de la DA en el córtex prefrontal en «NEUROCIENCIA DE LA RELIGIÓN (II): DOPAMINA, PENSAMIENTO MÁGICO Y RELIGIÓN».
10. «Los seres humanos están naturalmente inclinados hacia las creencias sobrenaturales. Muchos individuos altamente educados e inteligentes experimentan una poderosa sensación de que hay patrones, fuerzas, energías, y entidades que operan en el mundo que son negadas por la ciencia porque van más allá de los límites de los fenómenos naturales que entendemos actualmente. Más importantemente, tales experiencias no están sostenidas por un cuerpo de evidencia fiable, razón por la cual son sobrenaturales y no científicas. La inclinación o el sentido de que ello puede ser real es nuestro súper-sentido» (Hood 2009 p. X).
11. Por supuesto, el PM está sentado como una dimensión intrínseca de la esquizotipia (p.ej. Eckblad & Chapman 1983; Reynolds et al. 2000; Mohr et al. 2006; Unterrainer et al. 2011).
12. Empero, el mismo equipo de investigadores fracasó en otro estudio buscando la correlación DA/creencia paranormal (Palmer et al. 2007). Siendo ciertamente robusta la evidencia a favor, los autores discuten probables fallas metodológicas en el descarte de la predictibilidad/susceptibilidad de los participantes ante las tareas destinadas en el estudio. De hecho, más evidencia reciente muestra que la potenciación de dopamina en personas sanas aumenta sus atribuciones sociales paranoides (Barnby et al. 2019).
13. Todo esto está enmarcado en el aspecto evolutivo del aumento del tamaño de los grupos sociales durante las últimas fases de la hominización, incrementándose la competencia social y el neocórtex (p.ej. Dunbar 1992; Aiello & Dunbar 1993), haciendo aparecer así nuevos estresores o al menos maximizándolos y, mientras así emergiendo el cerebro social (p.ej. Dunbar & Shultz 2007), canalizando mecanismos y funciones hacia el manejo del estrés.
Resumen
«Si la magia es tan evidentemente falsa desde una perspectiva empírica, ¿por qué abraza tan tenazmente a la imaginación humana?» (Brown 1997)
Ciertamente, el cerebro humano no es una máquina de discernimiento analítico-empírico. Es decir, el pensamiento/conducta de tipo científico (p.ej. observar, elaborar teorías e hipótesis, realizar experimentos y evaluar la evidencia obtenida para llegar a conclusiones) es de «naturaleza innatural» (McCauley en Keil & Wilson 2000). Entonces, ¿puede ser que el llamado ‘pensamiento mágico’ sea una estrategia mental/conductual adaptativa? Dentro del marco del descubrimiento y estudio de aspectos cognitivos naturales estructurando a la religión y a la religiosidad, ¿es el pensamiento mágico, y no la religiosidad ni la religión, lo que con exactitud es intuitivo e innato? ¿Podría tener el pensamiento mágico una base biológica evolutiva? Para hacer consistentes estas cuestiones y poder plantear respuestas:- en la PARTE 2 de este largo ensayo, se dejó sentado que el pensamiento mágico y la religión son aspectos relacionados íntimamente pero independientes: parece inconcebible que exista religión y religiosidad (y también espiritualidad) sin pensamiento mágico, sin embargo éste sí existe (y persiste) sin religión; y adicionalmente, existe en prácticamente cualquier ámbito de la vida cotidiana y aún (persiste) en nichos socioculturales, así como en las mentes individuales, en principio no religiosos (p.ej. coexistencia cognitiva entre ciencia y pensamiento mágico: Legare et al. 2012);
- luego, más allá de su auténtica universalidad, persistencia y prevalencia, y más allá aún de producir errores de interpretación cognitiva (p.ej. en contraste con las explicaciones científicas), el pensamiento mágico parece estar motivado por profundos aspectos motivacionales como la supervivencia (p.ej. al intuir un potencial peligro ante lo que en realidad es información poco clara del entorno —como se mostró en el experimento de la caja «mágica» que podía explotar al introducir la mano), o la socialización (como en el caso de quienes creen que la mirada de los demás influye —mágicamente— en ellos, tal como si la percepción de las intenciones de los demás y su relación con el individuo estuviera hipertrofiada);
- consistentemente, hay una directa relación al menos demostrada psicológicamente, entre el pensamiento mágico, la ansiedad y el estrés —aquél parece un efecto regulatorio ante éstos dos, por lo que se espera la existencia de un sustrato neural que explique tal relación: como se vio, la dopaminergia involucrada es un fuerte indicio de tal sustrato. Esto nos introduce de lleno a una dimensión del pensamiento mágico aún no explorada por la ciencia: su neurobiología.
Y, habiendo tomado el concepto del psicólogo E. Subbotsky de que el PM implica que «los objetos físicos en el final receptivo de un proceso comunicativo tienen algún tipo de consciencia» (2010 p. 9 § 1 —libro completo disponible aquí), se concluye que el PM es simplemente la hipertrofia misma de la socialización humana. Lo que a su vez equivale al concepto del antropólogo S. Guthrie de que los humanos estamos innatamente capacitados para «tratar con el mundo en general como si fuera social y comunicativo» (2002 —una propuesta original de Walter Burkert). El trabajo de Subbotsky es sobre el pensamiento mágico, y el de Guthrie sobre el animismo y sus fundamentos filogenéticos: se trata entonces de lo mismo, conductual y cognitivamente, cuando nos referimos a ‘hiperagencia’, ‘pensamiento mágico’ y ‘animismo’. Así, estos términos refieren a un mismo proceso cognitivo, que en el ámbito académico se ha denominado «Hyperactive/hypersensitive agent-detection device (HADD)» [en español: dispositivo hiperactivo o hipersensitivo de detección de agente o de agencia] (Barrett 2004). Si el PM es parte misma, es decir natural, de la socialización, se predice la existencia de un sistema neurocognitivo modular y de carácter innato que le de soporte. Este sustrato debería formar parte del así llamado ‘cerebro social’, en sí mismo conformando la función de hiperagencia del sistema de Detección de Agencia, el cual es pues considerado el núcleo del cerebro social (Frith & Frith 2010).
Todo esto contrasta con la religiosidad (R), que no es un módulo cognitivo ni un patrón funcional específico, sino una compleja colección de varios aspectos (ideacionales/conductuales, intuitivos/racionales, individuales/grupales, genéticos/adquiridos y circunstanciales), que resultan no exclusivamente enfocados hacia la R misma sino que se trata de mecanismos generales de cohesión grupal e interacción social (p.ej. también identificables en la política o el deporte). Sin embargo, diversos autores y publicaciones recientes han dado más fuerza a una hipótesis adaptacionista de la religión y/o la R, que implicaría que toda-ella es un asunto de selección genética, en calidad de promotora (y supuestamente originadora) de la cohesión/cooperación social. Aunque aclarado ya en la PARTE 1 que no es el marco hipotético seguido aquí, nos introduciremos en esta tercera parte volviendo a tocar este tema, que resultará en un definitivo contraste entre el PM y la R.
2. La ‘naturalidad’ del pensamiento mágico y la ‘innaturalidad’ de la religiosidad: más indicios
2.1. ¿BASES GENÉTICAS DE LA RELIGIOSIDAD?
Recientemente, la elevada taza de fertilidad dentro de ciertas congregaciones religiosas (en EE.UU.) otra vez se muestra como la clave de su expansión: la diseminación de sus doctrinas depende más de la cantidad de hijos, que serán pues adoctrinados, que del proselitismo que se ejerza para ganar nuevos adeptos y mantenerlos adheridos (Scheitle et al. 2011). Este y otros datos estadísticos sobre la relación fertilidad/R son tomados como un indicativo de que la R tiene una base genética (ver «EXPLICANDO LA RELIGIÓN, CONFERENCIA EN BRISTOL 2010»), mientras que otros datos similares sugieren no es ella misma la causa de tal fertilidad (ver «¿POR QUÉ LOS AUSTRÍACOS RELIGIOSOS TIENEN MÁS HIJOS?»). Un más reciente análisis arroja que los ciclos ovulatorios de 275 mujeres estadounidenses, se asocian a fluctuaciones en su religiosidad (y postura política), en el sentido de que las solteras fueron menos religiosas (y menos políticamente conservadoras, que es lo mismo que más liberales) mientras ovulaban, o sea siendo más fértiles, y las mujeres con relaciones comprometidas (p.ej. casadas) fueron más religiosas (y menos liberales) cuando más fértiles (Durante et al. 2013). Los autores creen que el incremento de religiosidad (y conservadurismo) «pueden servir para disuadir a las mujeres casadas de engañar a su cónyuge», y aunque no exploran tal posibilidad (p. 9), en cambio parece más consistente que, «dado que la ovulación lleva a las mujeres a ser más abiertas a las relaciones sexuales de corto plazo, la ovulación puede alterar las actitudes religiosas y políticas de las mujeres para facilitar este tipo de relaciones. Dado que la apertura a las relaciones sexuales de corto plazo se asocia con la baja religiosidad [...] y las actitudes políticas más liberales [...], la ovulación puede llevar a las mujeres a ser menos religiosas y más liberales» (p. 2). Como se vio en la PARTE 2 y se verá más abajo sobre los marcadores neuroquímicos y hormonales de la empatía y el emparejamiento, hay pues sólidas razones para ver aquí la flecha de causalidad en sentido neurocognición→religión, y no al revés.
Anteriormente se señalaron otros datos estadísticos/socio-demográficos significativamente contradictorios, que no permiten pues concluir que la R tenga alguna base genética a su expreso servicio, en tanto que esta conclusión está ciertamente sesgada a valorar positivamente la R como necesariamente beneficiaria de la salud, o bien está sesgada cristianocéntricamente, como es el caso de los argumentos de M. Blume (p.ej. Blume 2011). Notoriamente, hay una especie de salto entre lo definido como R y la presunta base genética asociada: las correlaciones positivas obtenidas con gemelos y el grado de R, y p.ej. este estudio de Scheitle et al., tendrían que entenderse como evidencia de que un gen o varios genes controlarían la devoción, la afiliación, el conocimiento doctrinal, la creencia, la asistencia a un centro de culto y la espiritualidad, en conjunto, ya que todo esto mismo es lo que se define como R (aunque ciertamente no todos los elementos se presenten necesariamente a la vez). Véase que, como puede comprenderse haciendo un recorrido por nuestras publicaciones, tan solo la creencia afirmativa en «agentes supra-empíricos» (en palabras de Blume) es por sí misma un proceso cognitivo/emocional, estructurado así por «autorrepresentación, asociación emocional, recompensa y conductas dirigidas a metas», todo lo que, sobresalientemente, es un estado neural indistinguible de cualquier otra creencia afirmativa ordinaria (ver «CORRELATOS NEURALES DE LAS CREENCIAS RELIGIOSAS Y NO RELIGIOSAS»). Esto de por sí pone en seria duda que exista algún proceso neurocognitivo funcionando solo para la R, o algún estado cerebromental significativamente independiente (todo esto más bien va en apoyo de la teoría del subproducto), y luego veremos aún más al respecto. Entre tanto, además la convicción en las afirmaciones religiosas puede verse influida por un contexto fuertemente socio-afectivo, tal como una comunidad religiosa a la que se hace adepto un individuo (ver «¿POR QUÉ LA RELIGIÓN HACE A LA GENTE MÁS FELIZ?»).
Aún ¿cómo es que, un solo gen o una cantidad de genes, controlaría todo eso que es la R, cual patrón ideacional/conductual? La respuesta no es que los genes sí lo puedan hacer ya que, en efecto, sí hay patrones conductuales demostrados bajo influencia genética (p.ej. tan específicos como el miedo: Hariri et al. 2002; Lau et al. 2009; Inoue et al. 2011; o aún la prosocialidad:2 Knafo et al. 2007; Macdonald & Macdonald 2010; Striepens et al. 2011), sino que, evidentemente, la R no es un patrón tal: de hecho, la prosocialidad misma y el miedo subyacen a algunos de sus componentes (p.ej. ‘afiliación’), y, también evidente, la prosocialidad y el miedo en absoluto están exclusiva ni expresamente dedicados a la R, ni a la ‘afiliación’. Es ya inconcebible biológicamente, en tanto que Scheitle et al. muestran que incluso una doctrina correlaciona con más fuerza que otra, dentro de una misma religión (cristianismo), con la fórmula «predica-luego-engendra». Es decir, si es evidente que no podemos esperar encontrar genes regulando lo que no es sino un conjunto relativamente homogéneo de diversos aspectos (porque no necesariamente en todos los individuos se combinan todos los aspectos que integran la R, o estos mismos varían significantemente a través de las culturas y la historia, en contraste con el miedo o la prosocialidad), mucho menos aún cabe una «genética del evangelista» ni «del mormón», como si fuese si quiera concebible un contraste doctrinal a tal nivel biológico-genético. Analizando con más detenimiento el argumento de Blume, teniendo en cuenta la definición de R y los estudios en Ciencia Cognitiva de la Religión (incluyendo en su campo el adaptacionismo prosocial: p.ej. Wilson 2003), la conclusión es que estos datos no están sugiriendo que la R ni la religión, mucho menos aún alguna específica doctrina cristiana, esté determinada genéticamente.
P.ej. Blume (2011 p. 9) interpreta los resultados de Schjoedt et al. 2009, y Urgesi et al. 2010, como indicios de una biología adaptacionista de la R, debido en estos casos a la ‘neuro-modularidad’ que evidencian para la oración y la espiritualidad (o autotrascendencia), respectivamente: la ‘oración personal’ requiere las cortezas temporo-parietal, temporopolar y frontomedial; y la autotrascendencia recluta la corteza parietal inferior y la junción temporoparietal. Pero Blume pasa por alto que precisamente tales ‘neuro-modularidades’ respectivamente consisten en la ‘Teoría de la Mente’ (ToM: Gallagher & Frith 2003; Saxe1 & Powell 2006; Saxe 2010) y la ‘percepción espacial’ [autorreferencial] (Wilson & Keil 2001 pp. 784-787; el córtex parietal inferior como sustrato de la distinción yo/otros: ver «NEUROCIENCIA DE LA RELIGIÓN (VII): EL PAPEL DEL LÓBULO PARIETAL»; el córtex parietal posterior/superior/inferior como procesador egocéntrico del espacio: Galati et al. 2000; Weniger et al. 2009; Ciaramelli et al. 2010). Ambos aspectos tienen funcionalidad generalista, y están asociados (p.ej. en la interacción social —véanse las referencias sobre la ToM; o la prospección —véase sobre el Mental Time Travel), en lugar de parecer ‘módulos’ única y expresamente soportando la oración y la autotrascendencia, y no otros aspectos ideacionales y/o conductuales (como los ejemplificados). De hecho, Schjoedt et al. sostienen «que rezar a Dios es una experiencia intersubjetiva comparable a la ‘normal’ interacción interpersonal» (p. 205 § 8); mientras Urgesi et al. que «la reducción de la actividad neural en el córtex temporoparietal durante las experiencias espirituales puede reflejar un alterado sentido del propio cuerpo en el espacio» (p. 316 § 2). Otra vez: (en realidad) ambos estudios no sugieren reclusión neurocognitiva exclusiva al servicio de la R.
Téngase en cuenta que respecto a ambos, ToM (por los menos sus rudimentos) y percepción espacial, se consideran funciones filogenéticas (sobre ToM: cf. Herrmann et al. 2007; Call & Tomasello 2008; Schmelz et al. 2011; Krupenye & Call 2019; sobre percepción espacial: cf. Haun et al. 2006), e innatas, o mejor dicho, que si bien no están operativas al momento de nacer, su temprano surgimiento (cf. Poulin-Dubois et al. 2009) y desarrollo postnatal dependen de redes neurales cuya formación estaría contralada genéticamente por lo menos en aspectos críticos (como el reconocimiento facial respecto a la ToM), que es lo que se espera hallar en un cerebro adaptado para la socialización (véase como ilustración «Teoría de la Mente y neurodesarrollo»). Hay que elaborar un poco más este punto. Veamos, es capital recordar que el córtex temporomedial (p.ej. hipocampo), que contribuye a la representación espacial alocéntrica, es una región con baja influencia genética pero ambiental elevada; mientras que el córtex parietal, asociado al espacio egocéntrico, resulta más influido genéticamente (ver la sección «Sobre el cerebro y la influencia genética: relaciones con la cognición religiosa» de la PARTE 1). Adicionalmente, la interconexión entre las cortezas parietales izquierda/derecha también depende más del control genético, que ambiental, en comparación con las cortezas frontales, donde ocurre lo contrario respecto al cuerpo calloso (el grueso haz de fibras nerviosas interconectando ambos hemisferios cerebrales). Sería necesario señalar que tales redes neuronales aumentan o disminuyen su actividad e interconexión por neurotransmisión y neuromodulación, mediante una cantidad de moléculas cuya síntesis, cantidad y actividad sinérgica óptima, o típica, es controlada genética y epigenéticamente, conformando «valores» fijados evolutivamente acordando con el concepto de Edelman & Tononi (2000 pp. 30-33, 62-65). Es decir, es dudoso que la historia de la selección natural de los genotipos y fenotipos implicados en estos procesos (aunque aún solo estemos empezando a identificarlos específicamente) haya sido escrita por la cronológicamente muy reciente aparición de la R en el escenario de la encefalización homínida. Llegamos así al punto del descubrimiento de las bases neurales de la religión y la R: los estudios al respecto muestran que su variación entre individuos correlaciona con las variaciones neuroanatómicas y funcionales dedicadas a la cognición social (Kapogiannis et al. 2009a; 2009b), que como notan estos autores, es la misma clase de diferencia p.ej. entre músicos y no-músicos.
2.2. LA RELIGIOSIDAD EN EL CEREBRO: UN SUBPRODUCTO Y NO UNA ADAPTACIÓN
En el estudio de Kapogiannis et al. (2009a), se analizó el córtex cerebral de 40 individuos adultos en cuanto a estructura y funcionalidad, mientras que un test psicológico daba cuenta de su religiosidad (R). Así, «experimentar una íntima relación con Dios» (manifiesto en practicar la oración y la participación religiosa), correlaciona con el grosor del córtex temporal lateral (giro medio derecho), que en cuestión ya hace varios años ha sido identificado como un «lugar» del cerebro para la religión,3 y que, en lo que se refiere a Dios en cuanto a ‘sus intenciones’, ‘sus emociones’ y ‘su relación’ con el individuo, el mediador es la ToM. Esta región contribuye al autorreconocimiento y la evaluación de las relaciones íntimas vinculadas a la distinción yo/otro, y su variabilidad estructural desproporcionada, p.ej. hacia menor o mayor tamaño, se asocia respectivamente a la esquizofrenia o la obsesión. En base a esto, los autores especulan con que el rango de variación volumétrica de esta región sostenga un espectro psicológico, que va desde el normal-mayor volumen cortical y la conducta ritual, el normal-bajo y la irreligiosidad, y ya el volumen patológicamente bajo y la esquizofrenia, que implica pues un trastorno de la distinción yo/otro (yo/Dios) (ibid. pp. 4-5). Por otro lado, «experimentar miedo a la ira de Dios» depende del córtex orbitofrontal (CFO en adelante), también asociado a la ToM en cuanto a aspectos emocionales (empatía) y su regulación; y el precuneus, asociado al «yo» y a la recuperación de recuerdos (Mental Time Travel —MTT), registrado en la circunstancia de relación de Dios con el yo (p.ej. en la oración). Nuevamente, el poco volumen cortical en ambas regiones puede predisponer a un miedo a Dios, al estar comprometidas así las funciones de regulación emocional y relación comunicativa con ‘él’; mientras que, inversamente, es posible que el mayor volumen del precuneus correlacione con poner énfasis en las experiencias mundanas en lugar de las internas, facilitando así una postura no-religiosa (ibid. p. 5). Finalmente, este estudio no encontró ninguna diferencia cortical volumétrica, que sea significante, correlacionando con la educación religiosa temprana (infancia) recibida por los individuos; lo que indica que, en tanto no hallada ninguna modificación especial ni importante en el cerebro asociada a la religiosidad, «cualquier otro efecto inicial de la educación religiosa puede ser disipada por las experiencias posteriores de la vida». Los autores concluyen que:
«Las áreas del cerebro identificadas en este y los estudios en paralelo de fMRI no son exclusivas del procesamiento de la religión, sino que juegan un papel importante en la cognición social. Esto implica que las creencias y la conducta religiosa no surgieron como adaptaciones evolutivas sui generis, sino como una extensión (algunos dirían “subproducto”) de la cognición y el comportamiento social. Además, el estudio actual sugiere que la evolución de ciertas áreas que desarrollaron la comprensión y la empatía hacia otros seres humanos (precuneus, CFO, temporopolar/lateral) puede, al mismo tiempo, haber permitido una relación con un agente sobrenatural (Dios) basada en la intimidad en lugar del miedo.»
Un estudio más reciente, realizado con 268 adultos mayores, encontró una relación, independiente de factores psicosociales o demográficos, del tamaño del hipocampo con los cambios importantes en las creencias religiosas (Owen et al. 2011). Una mayor atrofia hipocampal se encontró en los individuos que habían vivido un «renacimiento» o una experiencia religiosa, y en base a ello los autores postulan que esto se debería al «impacto del estrés acumulativo» (p. 2). Es decir, «mientras algunas variables religiosas se han asociado a la salud mental, otros factores religiosos pueden ser una fuente de estrés» (p. 3); y lo segundo implica un impacto neurobiológico pernicioso para el hipocampo, una conocida región cerebral especialmente sensible al estrés, incluyendo p.ej. acumulación de glucocorticoides y muerte neuronal. Estas circunstancias religiosas pertenecen en realidad a un fenómeno minoritario que puede ocasionar estrés en el individuo, paradójicamente «aún si la experiencia fue subjetivamente positiva», debido a que «las experiencias no interpretadas con facilidad dentro de un existente marco cognitivo o de creencias religiosas han sido mostradas como perjudiciales para el bienestar subjetivo», y «pueden también quebrar los lazos sociales» (p. 3). Los autores notan que la atrofia hipocampal está involucrada en diversos cuadros clínicos como depresión, demencia o Alzheimer, que se relacionan precisamente al estrés acumulativo, y finalmente concluyen que esta atrofia resulta de la influencia de factores religiosos. Sin embargo, A. Newberg (Scientific American 2011), uno de los representantes de la así llamada «neuroteología», comenta sobre este estudio y dice que, cambiando la flecha direccional de causalidad, puede «que las personas con menor volumen hipocampal tienen más probabilidad de tener atributos religiosos específicos», y que «los factores que condujeron a los acontecimientos que cambian la vida son importantes y no sólo la propia experiencia» (religiosa). «Dado que la atrofia cerebral refleja todo lo que le sucede a una persona hasta ese momento, uno no puede concluir que la experiencia más intensa fue, de hecho, lo que dio lugar a la atrofia cerebral».Newberg acierta en subrayar la falta de correlación del estrés con la atrofia hipocampal en este estudio (que los propios autores notan en su artículo como una de las limitaciones), cuando es en cuestión la causa postulada. De hecho cabe aún la posibilidad de que, a pesar que ambivalentemente la religiosidad es estresora y anti-estrés, en estos individuos la religión haya sido una protección contra el estrés causado por factores no religiosos. Ahora bien, si la hipótesis de la R como subproducto es correcta, entonces esto es más evidencia de que no hay una determinación biológica, sino que existen ciertos rasgos neurocognitivos (p.ej. mayor/menor volumen hipocampal) que se modulan según las experiencias y el aprendizaje. De hecho se ha mostrado que el hipocampo puede cambiar físico-estructuralmente (el proceso se llama neuroplasticidad) tras un lapso tan corto como 2 horas de aprendizaje (Sagi et al. 2012; el mismo estudio realizó una replicación en ratas). Entre tanto, se conocen las funciones hipocampales como memoria, Mental Time Travel, procesos emocionales, aprendizaje o navegación espacial, todo lo cual es también conocido por estructurar las experiencias religiosas (un aspecto importante de la R).
Así, se ha definido un «marcador límbico» (‘Sistema Límbico’ es un término clásico para agrupar ciertas estructuras cerebrales que incluyen al hipocampo, la amígdala y también el córtex frontal orbital o CFO) para tales experiencias: «la actividad límbica puede marcar las siguientes experiencias: 1) despersonalizada o desrealizada, 2) de crucial importancia autorreferente, 3) armoniosa-indicativa de una conexión o unidad entre elementos dispares, y 4) de éxtasis, profundamente alegre. Esta actividad límbica subyace a ciertos ataques epilépticos con auras, las experiencias cercanas a la muerte, y las experiencias místicas y religiosas de los individuos normales» (Saver & Rabin 1997 p. 507 § 4). Sabemos que numerosas circunstancias como fiebre, ensueño, danza, falta de oxígeno, enteógenos, orgasmo, etc., y en individuos religiosos o no, pueden conducir a cualquiera de esas experiencias, aisladas o concurrentes. Entonces, los contenidos vivenciales de los ‘estados alterados de consciencia’ no son necesariamente interpretables como experiencias religiosas (si bien es cierto que el marco de creencias y sesgos de quien tiene la experiencia determinará la interpretación narrativa y la lógica explicativa de la misma); simplemente, toda esta fenomenología puede tratarse de la supresión del estado de vigilia (cf. Vaitl et al. 2005). Eso es lo que sugiere la asociación entre la funcionalidad cerebral, las experiencias religiosas, la autotrascendencia y los estados alterados de consciencia, en lo que solo recientemente se ha descubierto que es importante un sistema de regiones cerebrales interconectadas denominada Red por Defecto (RpD; en inglés Default-Mode Network).
2.3. LA IDEACIÓN MÁGICA: MODO DEFAULT DEL CEREBRO
Esta Red por Defecto o RpD (ver Fig. 1 abajo, ver aquí también) y su actividad espontánea e inconsciente, parece reflejar una organización básica del cerebro. Se ha visto que las tareas conscientes cuando evocadas (en contraste con las tareas que se realizan fuera de los experimentos) consumen notoriamente menos recursos (suponen menos funcionalidad estructural y por ende menor trabajo sináptico y gasto metabólico), y como se sabe, cuando cesan, p.ej. si no se está haciendo/pensando nada, en cambio aumenta la actividad de la RpD (Boly et al. 2008 p. 2). No está claro cuál es la función exacta de esta red, aunque debe señalarse que se ha mostrado asociada a procesos introspectivos y autorreferenciales, y parece un organizador funcional de la consciencia. El umbral óptimo de ésta es un balance entre el estado de vigilia (de correlato subcortical: hipotálamo, tronco encefálico, formación reticular, prosencéfalo basal —regulando además los estados vigilia/sueño) y la consciencia per se, es decir sus contenidos (de correlato cortical: frontoparietal, y subcortical: tálamo), y ésta última se subdivide en autoconsciencia y consciencia de lo externo, que en cambio están inversamente correlacionadas. La consciencia de lo externo involucra las regiones laterales frontal/parietal, mientras que la autoconsciencia es la propia RpD: cortezas cingulada posterior, precuneus, frontomedial y temporoparietal. Desconexiones dentro de estas redes tálamo-cortical (consciencia en general) y cortico-cortical (autoconsciencia/consciencia de lo externo) han sido reportadas en estados alterados de consciencia como el sueño y la anestesia, mientras que la recuperación de un estado vegetativo implica la restauración de tales redes (ibid. pp. 4-5; p. 15 Fig. 2, ver abajo, cf. p. 16 Fig. 3).

Fig. 1. Comparación (A) del tracto conectivo (materia blanca) perisilviano, (B) RpD (Red por Defecto), (C) el MTT (prospección), (D) la ToM (Teoría de la Mente), (E) la agencia, y (F) el espacio extrapersonal. Se trata de sistemas diferentes pero todos compartiendo el «Núcleo de Agencia», asociados así a la hiperagencia —y por tanto al pensamiento mágico o al animismo. Pinchar en la imagen para ampliar. (Ver también aquí, y aquí.)

Fig. 2. «(Izquierda) La consciencia tiene dos mayores componentes: la activación [‘arousal’: ver definición en español], o el nivel de consciencia, y la consciencia [‘awareness’], correspondientes a los contenidos de la consciencia en sí mismos. La activación [‘arousal’] y la consciencia están por lo general positivamente correlacionadas. Sin embargo, involucran a diferentes estructuras cerebrales. La activación [‘arousal’] consiste en la actividad de las estructuras subcorticales abarcando el tronco encefálico, la formación reticular, el hipotálamo y el prosencéfalo basal. La consciencia [‘awareness’] se relaciona con la actividad de un amplio conjunto de áreas asociativas fronto-parietales, lateral y medialmente. (Derecha) La consciencia [‘awareness’] puede a su vez dividirse en dos mayores componentes: autoconsciencia y consciencia de lo externo. […] La autoconsciencia abarca las cortezas cingulada posterior/precuneus, frontomedial, y la junción temporoparietal bilateralmente. La consciencia de lo externo abarca las cortezas laterales frontal y parietal». (Adaptado de Boly et al. 2008 p. 15 Fig. 2.)
Pienso que la sugerencia hecha es particularmente fuerte como parte de una neurociencia de las intuiciones hiperagentivas (teleológicas, animistas, mágicas), y la elaboración deliberada de éstas bajo la forma de creencias de contenido mágico y/o religioso. La RpD comprende subsistemas intuitivos como la Detección de Agencia, fundamental aquí, mientras que la red frontoparietal lateral comprende regiones que la revisten de imaginación, creatividad y narrativa lógica, planificando y ejecutando las acciones pertinentes. Hay adicionalmente un parecido entre esta división neurocognitiva relacionada inversamente, y la teoría del Procesamiento Dual, que propone que la actividad mental se compone paralelamente del Sistema 1 intuitivo y automático, y otro Sistema 2 analítico y lento. Este modelo recientemente está echando luz a la estructura cognitiva de la creencia (y descrecencia) en Dios (ver aquí, y aquí). Ciertamente los correlatos neurales asociados al Sistema 1 no se corresponden con la RpD, mientras el Sistema 2 se basa en una red frontoparietal similar a la consciencia de lo externo. Sin embargo, parece evidente que las fuentes intuitivas del Sistema 1 estan contenidas en la RpD, y aún otras redes descubiertas que también están activas mientras se está en reposo (Broyd et al. 2009 p. 4).
En consonancia con todo esto, Baumard & Boyer (2013) modelan las creencias religiosas como el producto conjunto de las intuiciones, es decir «la salida de sistemas cognitivos especializados», y las reflexiones sobre éstas o «metarepresentaciones de las intuiciones». Aunque este modelo hace en efecto tal distinción, los autores nos dicen que no necesariamente se trata de dos amplios sistemas: «las intuiciones y las reflexiones son probablemente producidas por un gran número de sistemas de dominio específico. Además, las reflexiones no son paralelas a las intuiciones, como implican algunos modelos de procesamiento dual. Las reflexiones son disparadas y limitadas por el contenido específico de las intuiciones» (p. 3). Se trata de un modelo de múltiples procesos jerárquicos como explicación de lo que es una creencia religiosa, que resta demostrar neurocientíficamente, pero del que ya conocemos fundamentales aspectos intuitivos como la detección de movimiento biológico o el reconocimiento facial, y otros aspectos de carácter reflexivo como las funciones prefrontales dorsolaterales de elaboración de narrativa explicativa. Notoriamente, el funcionamiento alterado de la RpD, natural y artificialmente inducido, constituye los estados alterados de consciencia, manifiesto en lo que respecta a hiperagencia (~ agencia sobrenatural) y los contenidos mágicos en las llamadas experiencias religiosas. Esto es consistente con que la RpD está constituida por agencia, ToM, MTT y mapeo espacial, lo cual es también soportado por la implicación de la RpD en los estados alterados de consciencia de la epilepsia (Danielson et al. 2011), en la esquizofrenia (Öngür et al. 2010), o en la experiencia de la meditación (Brewer et al. 2011), todos estados mentales caracterizados por alteración de la agencia y sus contenidos mágicos.
Otro hecho crítico en tanto la RpD está formada por agencia, ToM, MTT y mapeo espacial, es que si el hombre es un primate híper o ultrasocial, esta es exactamente la estructura anatómica y la organización funcional que se espera encontrar en su cerebro. Ahora bien, si comparamos los sustratos neuroanatómicos de la autoconsciencia (RpD), con la consciencia de lo externo (lateral frontoparietal), notaremos que en realidad sus partes posteriores en el cerebro corresponden similarmente al córtex parietal. Por supuesto, el córtex parietal tiene importantes subdivisiones corticales/funcionales: las regiones superior/posterior enfocadas a la orientación y el mapeo espacial; somatosensorialidad (mapeo del cuerpo) hacia la frontera con el lóbulo frontal y la región motora; la lateral inferior hacia la frontera con el córtex temporal sirviendo a la distinción yo/otros y la semántica (área de Wernicke); el surco intraparietal (dividiendo las regiones superior/inferior) implicado en la dirección de la mirada, la detección de intención y el sentido numérico; y finalmente la zona medial de la región superior en el surco interhemisférico o precuneus, asociado al sentido autorreferencial, y que aparece pues como un importante nódulo funcional de la RpD y la red frontoparietal de la imaginación y la consciencia de lo externo.
Estas subdivisiones parietales están interconectadas entre sí, y es por ello que funcionalmente este lóbulo es el foco de la sensación del cuerpo, de la percepción del espacio que le rodea y de su ubicación en él, de la preparación de la intención en este espacio y la capacidad de cálculo numérico/geométrico (p.ej. para dirigirse hacia un punto o coger un objeto), y, probablemente como producto de la expansión temporal-parietal, la sensación de propiedad sobre el cuerpo y el sentido del ‘yo’ como adaptación al crecimiento del grupo de individuos (prácticamente todo a lo que subsirve la afinación de las demás funciones parietales respecto a otros primates: p.ej. exploración, fabricación de herramientas, organización social). Consistentemente, el precuneus «es relativamente más grande en los seres humanos que en los primates no humanos y otros animales, y es una de las últimas regiones en mielinizar durante el desarrollo» (Schlegel et al. 2013 p. 4), y es de hecho, junto con el cerebelo, la zona más expandida durante la globuralización evolutiva del cerebro humano (Bruner et al. 2017; Neubauer et al. 2018). En consecuencia, ambas redes diferenciadas respectivamente para la autoconsciencia y la consciencia de lo externo, evidentemente comparten un mismo sistema posterior, el córtex parietal en general (y más específicamente el precuneus), que es ciertamente espacio-egocéntrico involucrando sus regiones superiores y laterales (Galati et al. 2000; Weniger et al. 2009; Ciaramelli et al. 2010), tal como se ha mostrado líneas atrás mientras se discutían los estudios sobre la oración personalizada y la autotrascendencia.
Nuevamente, como analizamos aquí sobre la RpD, la red frontoparietal, el córtex temporal medial como el hipocampo y regiones adyacentes (córtex fusiforme, frontoventral, parahipocampo, amígdala —todas también implicadas en la religión), tampoco ningún dato sugiere que trabajen exclusivamente para la religión. Parece que más bien la religión y sus ideaciones extraordinarias son de hecho un subproducto de la funcionalidad interconectada de estas redes. La ‘cognición religiosa’ se entiende así más claramente, en principio, como el tipo de ideación que se espera encontrar como resultado de tal organización neurocognitiva.
3. Religiosidad y prosocialidad: en realidad nada inusual
3.1. LA MATRIZ BIOLÓGICA (Y NO RELIGIOSA) DE LA COOPERACIÓN SOCIAL
Otro fundamento del adaptacionismo aquí es la observación de ciertas conductas, como la ‘señalización costosa’ —p.ej. rituales extremadamente estresantes, dirigidos a la cohesión/cooperación social (cf. Norenzayan & Shariff 2008). Ello, junto con el otro tipo de cohesivo social como el emparejamiento (y la fertilidad implicada), al cual ya nos remitió Blume, él supone que no causaría eventualmente (culturalmente) el surgimiento de la R, sino que ésta misma habría promovido la selección genética de tales conductas. Sin embargo Norenzayan & Shariff (2008), citados por Blume, concluyen que «aunque las religiones siguen siendo facilitadores de gran alcance de la prosocialidad en grupos grandes [de seres humanos], no son los únicos. La difusión cultural de instituciones seculares confiables, tales como los tribunales, las autoridades policiales, y los mecanismos eficaces para hacer cumplir los compromisos, aunque históricamente recientes, han cambiado el curso de la prosocialidad humana» (p. 62 § 4). Por supuesto, podrá argumentarse que aún tales instituciones seculares tienen en realidad fundamentos religiosos, pero esto no sería evidencia de una biología de la R, sino que la cooperación es más bien un aspecto biológico generalista (cf. Nowak & Highfield 2011; Krakauer 2011), en vista de que los compromisos ritualizados (y más o menos sacralizados) son parte integral de otras esferas sociales, como la política o bien cualquier agrupación con algún tipo de ‘identidad’ y ‘sentido de pertenencia’ (cf. Tajfel 2010 p. 2; Leary & Tangney 2005 p. 462), todo lo cual remite a motivaciones independientes de la R. Por último, de acuerdo con Boyer & Petersen (2011), las instituciones en las diversas culturas tienen rasgos compartidos (los autores abordan el matrimonio, la moral y el castigo del crimen, y el intercambio de bienes), que responden en cuanto a su diseño y estabilidad a las propias disposiciones cognitivas humanas.
Retomando la sección «La interacción social: estresora, a la vez que reguladora del estrés»: somos una especie hipersocial, o aún ultrasocial, implicando específicamente que los grupos sociales humanos pueden ser enormemente sofisticados, con la interacción y la competencia entre individuos (de hecho el concepto ‘Inteligencia Maquiavélica’, la capacidad de manipular a otros, es equivalente a ‘Inteligencia Social’) siendo potencialmente estresora y ansiogénica, por tanto también somos una especie «súper-cooperadora» y «pan-hiperagentiva» (p.ej. una enorme variedad de señales sociales debe ser no solo meramente identificable con rapidez, sino además, anticipada y proyectada coordinadamente) cuales dispositivos de regulación cognitiva/emocional en tal contexto. Un estudio reciente muestra por vez primera que el córtex orbitofrontal (CFO) tiene subdivisiones que correlacionan con aspectos específicos de la cognición social, aunque disociados, p.ej. el giro orbital izquierdo y la ‘Inteligencia Maquiavélica’, y el giro orbital derecho medio y el rasgo de personalidad ‘amabilidad’ (Nestor et al. 2012). Esta última refiere a la conducta prosocial, mientras el ‘maquiavelismo’ es lo opuesto en el Modelo de los Cinco Grandes rasgos de la personalidad, de hecho correlacionado al narcisismo y la psicopatía (Muris et al. 2017).
Se ha sugerido antes que esto se trata de un auténtico sustrato evolutivo-adaptativo, conformado por profundos ejes de combinaciones estructurales/funcionales neural, endocrino, gonadal e inmunológico. El maquiavelismo y la súper-cooperación humana son a la luz del evolucionismo un directo resultado del desarrollo del neocórtex en los mamíferos superiores y sobretodo en los primates, que como agregado neuroanatómico reciente muestra una funcionalidad relativamente bidireccional. Es decir, el ‘cerebro social’ humano está poderosamente influido por un complejo sistema de control neuro-endocrino de la conducta cooperativa, básicamente oxitocina y vasopresina, pero según los estímulos externos también tiene cierta capacidad de influencia sobre tal sistema (Soares et al. 2010), mientras que la testosterona está vinculada a la competencia y la conducta agresiva (Archer 2006; Victoroff et al. 2011). Este mismo sistema que sostiene nuestra conducta social conforma la respuesta de estrés e impacta en diversos sistemas orgánicos, como visto en la PARTE 2. Por otro lado la agencia, que Guthrie (2002) argumenta como extendida en el resto de animales, igualmente está asociada a tal sistema, hecho evidenciado porque el PM, que no es sino hiperagencia, tiene directa relación con el estrés, neurobiológicamente hablando mediante una vía frontomedial←→límbica←→LHPA, o con el reconocimiento facial-emocional mediante otra vía amigdalar←→LHPA (debe recordarse que la hiperagencia —la atribución de animación a los estímulos que no están animados— en sí misma consiste en la detección de movimiento biológico, y el reconocimiento de ojos/caras, que implican al Sistema Límbico). La prosocialidad está bajo influencia genética, como se decía más arriba, p.ej. respecto al gen receptor de vasopresina (Knafo et al. 2007), o el gen receptor de oxitocina (Striepens et al. 2011).
Ya en la PARTE 2 se mostró como la socialización está mediada neuroquímicamente por la interacción entre estas moléculas y otras como dopamina y serotonina. Se mostró que una y otra vez la dopamina se halla jugando un papel importante en la cognición social y el PM, que por lo visto bajo su modulación quedan relativamente ‘integrados’ a nivel funcional (además de traslaparse estructuralmente). Un significativo estudio citado allí, Domes et al. (2007), mostró que la aplicación intranasal de oxitocina «es suficiente para causar un sustancial incremento» en la inferencia de estados mentales, a través de la observación de la región de los ojos, en 20 de 30 individuos. Para esto se empleó el «Reading the Mind in the Eyes Test» (Baron-Cohen et al. 2001 —literalmente leyendo la mente en los ojos). En tanto que la amígdala es parte importante del reconocimiento facial/ocular, o que en ella in vitro la oxitocina estimula procesos inhibitorios, ésta «puede haber modulado la reactividad emocional de la amígdala» y/u otras regiones de la expectativa de recompensa como el núcleo accumbens (Domes et al. 2007 pp. 2-3). Oxitocina y dopamina interactúan para elaborar el output de la inferencia mentalística: es evidente que la hiperagencia dependerá de su modulación conjunta, pero, mientras se desconocen aún los procesos neurofisiológicos y neurogenéticos exactos, ya se han visto p.ej. importantes avances respecto a la dopamina y el PM. Mientras tanto, la testosterona inhibe la mentalización (Chapman et al. 2006; Baron-Cohen et al. 2011).
Oxitocina y dopamina se combinan para regular el apego, e interesantemente, el data acumulado indica sobre ello no solo bajo la forma de prosocialidad, es decir la aproximación y el enlazamiento empático hacia los congéneres sociales, sino también hacia los estímulos y las recompensas abstractas, distantes en el tiempo o ya imposibles factualmente. Es aquí cuando entra en juego como pegamento social la mentalización-hiperagencia y el PM con su amortiguamiento del estrés. Antes citábamos a Haidt (2011) para señalar que esto hace única la socialización humana entre los animales sociales. Enfocando esto evolutivamente, la fuerte cohesión social del Homo sapiens, entre otras capacidades cognitivas indisociables de ello como la fabricación de herramientas y el lenguaje (sobre las que recientemente se ha mostrado a nivel neural un posible origen co-evolutivo: Uomini & Meyer 2013), le ha permitido triunfar en diversos ambientes, superando mediante la organización coordinada el acecho de depredadores o la escasez de alimentos. Sin embargo alcanzar tal elevada complejidad en socialización implicó trasladar la ansiedad y el estrés a su campo, un fenómeno que no es exclusivamente humano sino que se observa ya en monos: «a diferencia de muchos animales cuya experiencia de estrés suele ser repentina y fuerte (p.ej. escapar de un predador al ataque), los babuinos del Serengeti son relativamente libres de la depredación y tienen un suministro abundante de alimentos por lo que sus principales estresores son de naturaleza social y son relativamente crónicos. Esta forma de estrés crónico es similar a los tipos de estresores a los que están expuestos los seres humanos» (Gosling & Harley en Corr & Matthews 2009 p. 282, aquí o aquí). Por lo visto hay una asociación neurobiológica y evolutiva entre el aumento del grupo social, el estrés social, la conducta prosocial y la formación de lazos sociales de tipo abstracto y trascendente, y esto mismo implica el desarrollo del PM.

3.2. RELIGIOSIDAD Y VIGILANCIA SOBRENATURAL: EL COMPONENTE CLAVE ES LA HIPERAGENCIA
La mecánica de la prosocialidad incluye aún otro aspecto, también tomado a favor de la R como adaptación biológica: el castigo sobrenatural. Esto es, «un miedo a la detección y el castigo “sobrenatural” sobre las acciones egoístas, que aumenta la aptitud darwiniana mediante la reducción de los costos de las transgresiones a las normas del “mundo real”» (Johnson 2011 p. 78 Fig. 1). Ya que esto es directamente lo que constituye la creencia en un dios que castiga el engaño (y como consecuencia habría mayor éxito reproductivo de los cooperadores en detrimento de los polizontes),4 es asumido que entonces la R estaría bajo control selectivo genético. Aunque a priori, nuevamente, no podría haber «una selección genética» de todo el sistema de ideas y conductas, privado y público, vinculado a ciertas creencias específicas y explícitas aquí implicadas (p.ej. ‘Dios castiga la mentira’), hay que detenerse en este asunto. Dando por sentado que ‘el hombre es un primate híper o ultrasocial’, intrínseco a esto son las siguientes circunstancias:
«Como las sociedades se expanden en tamaño, las relaciones sociales se vuelven cada vez más anónimas; el anonimato, eventualmente, hace que sea más difícil monitorear y castigar los fraudes y las conductas de no-cooperación, y como resultado los niveles de confianza se desploman y los polizontes imperan. Ante la falta de un monitoreo social exitoso, las sociedades colapsan [Dunbar 2003; Henrich 2006; Roes & Raymond 2003]. La histórica externalización del monitoreo social humano (en todas sus limitaciones) a la creencia generalizada en agentes omniscientes y moralmente comprometidos, podría haber aumentado enormemente la población de personas confiables de no cometer fraude, actuar como polizontes, u otra manera de transgredir las normas morales establecidas [Bering 2006; Johnson & Krüeger 2004; Roes & Raymond 2003].» (Shariff & Norenzayan 2011 p. 93 § 1)
Varios estudios muestran que «los grandes grupos pueden tener más polizontes que los pequeños grupos, pero ellos posiblemente tienen también más cooperadores»; mientras que el castigo como definitivo garantizador de la cooperación parece un «universal humano», a su vez que el aprendizaje y el contexto cultural influyen en ambos (Gächter & Herrmann 2009). Se ha encontrado que el castigo de polizontes es un aspecto uniforme transculturalmente, en contraste con las variaciones en los niveles de cooperación: «el castigo estabilizó la cooperación en todas partes» (ibid. p. 800 § 3). De hecho, de acuerdo con Johnson & Krüeger (2004), la cooperación social es persistente a pesar de que supuestamente no debería funcionar, p.ej. entre extraños no relacionados, pero «la cooperación en los grupos de la vida real se rompe (a pesar de que las mayores recompensas deben hacer cooperar a todos), a menos que se castigue a los tramposos»; y sin embargo, el propio castigo es un proceso costoso, y de ello pueden aún surgir «polizontes de segundo orden» (individuos que cooperan con el bien público pero que desisten de contribuir al castigo), y además, esto significa que «mientras el castigo parece ser necesario, no hay incentivo para quien lo realiza». Sin embargo hay evidencia de neuroimágen de que este castigo altruista es placentero: implica la activación del estriado (dopaminergia/recompensa), que incluso aumenta si es mayor el costo asumido al castigar (De Quervain et al. 2004); y se ha mostrado que los individuos asumen los mayores costos de castigo en la medida que aumenta la presión de competencia intergrupal (Sääksvuori et al. 2011) —esto también sugiere expectativa de recompensa de por medio, y se puede especular con que si bien no hay un incentivo para quien aplica el castigo, y sabiendo que la dopamina regula las recompensas abstractas y/o contrafactuales, entonces el incentivo pudiera ser igualmente percibido o esperado como algo abstracto, se trate del bien del grupo hacia futuro, o algún otro tipo de recompensa trascendental (p.ej. después de la muerte, en la otra vida).Entre tanto, las soluciones para el dilema de cómo mantener el castigo son: una institución externa que administre el castigo, que el castigo per se no sea costoso, y que tanto los polizontes como quienes desisten de castigarlos sean castigados; pero resulta que ninguna alternativa es del todo efectiva (Johnson & Krüeger 2004 p. 162). Luego, hay evidencia empírica de que el fraude del polizonte afecte emocionalmente (cause rabia) a quienes desisten de castigar, de tal manera que los lleve a asumir el costo de realizar el castigo; pero para esto fue necesario el anonimato en el experimento (Fehr & Gächter 2002). Johnson & Krüeger (2004) nos recuerdan que en la vida real pueden ser directamente confrontados los castigadores, lo que introduce el nuevo problema de que surjan represalias contra ellos. Por lo tanto, otra alternativa que toma forma es que la religión, con sus creencias en agentes sobrenaturales omnipresentes monitoreando la conducta, y castigando el egoísmo, haya funcionado como una primera institución de recompensa y castigo. Recuérdese que se habla de sociedades de un gran número de individuos ciertamente desconocidos entre sí, donde por lo tanto el peligro de fisión es mayor, y que ciertamente enfrentan constantes problemas de subsistencia (p.ej. la lucha por recursos, y la propia competencia con otras sociedades que ello implica), que para superarlos necesitan una efectiva cooperación masiva —hay en efecto una correlación entre el tamaño de la sociedad y la creencia en dioses moralizantes: «las sociedades confrontadas a las amenazas recurrentes a su existencia es probable que se caractericen por creencias en tales dioses» (Roes & Raymond 2003). Estas circunstancias problemáticas impactan negativamente en la sensación de control, son por tanto causalmente críticas para disparar la hiperagencia/PM (recordemos ahora la amplia evidencia que vincula PM y falta de control), y al parecer tienen una ubicación concreta en la (pre)historia: la Revolución Neolítica.
«Sólo el 23.9% de las 427 culturas en la base de datos intercultural reconoce un dios que está activo en los asuntos humanos, y es específicamente favorable a la moralidad humana. Las religiones con estos dioses son, de hecho, peculiares [...] la evidencia disponible sugiere la posibilidad de que la expansión de los rituales habituales y la construcción de arquitectura monumental religiosa significativa emergió junto con el aumento de tamaño de la sociedad, la complejidad política y la dependencia de la agricultura», como en las excavaciones neolíticas de Çatalhöyük (9500 años de antigüedad) y Göbekli Tepe (11000 años) (Norenzayan et al. 2012 p. 14). Respecto a los tiempos de registros escritos, el libro The Evolution Of God (Wright 2009) «proporciona un resumen útil de evidencia textual que revela la evolución gradual del Dios Abrahámico de un más bien limitado y caprichoso dios tribal de la guerra —un subordinado en el panteón— a la deidad unitaria, suprema, moralizante de dos de las mayores comunidades religiosas del mundo». Todo indica que la R juega un papel clave en la expansión de las comunidades prosociales desde hace unos 12000 años (ibid. pp. 14-15). Por lo tanto la historia, la arqueología, y la etnografía tampoco soportan una hipótesis adaptacionista de la religión, sino que señalan el más bien reciente surgimiento de esta como un promotor de la prosocialidad. De hecho las doctrinas de divinidades moralizantes toman su forma actual en la así llamada Era Axial, hace menos de 3000 años; y también, como las instituciones morales, las religiones moralizantes y sus notorias similaridades transculturales pueden ser mejor explicadas por una predisposición intuitiva, hacia la «proporcionalidad» entre acto y recompensa, pecado y penitencia, y la formulación de la Regla de Oro (Baumard & Boyer 2013).
Por cierto, Baumard & Boyer discuten brevemente sobre si las religiones moralizantes reforzaron las grandes sociedades (ibid. p. 276 Box 2. Did moral religions bolster large-scale societies?). Respecto a la hipótesis de Norenzayan et al. notan que «los dioses de la antigüedad no estaban modelados generalmente como interesados en las conductas morales o prosociales de las personas», y prosiguen ilustrándonos con ejemplos de exitosos imperios con dioses «sorprendentemente amorales» (Sumeria, Grecia, Roma, Mesoamérica, Inca). «Las religiones moralizantes tardías podrían no haber sido el disparo mágico que hizo posible las sociedades a gran escala, debido a que aparecieron mucho después de las primeras ciudades y reinos, 5000-10000 años más tarde.» Luego discuten la simultaneidad histórica de la emergencia de las relgiones moralizantes en los tres focos de la Revolución Neolítica (Mesopotamia-Grecia, India y China) (ibid. p. 277 Box 3. Why did moral religions emerge at the same time?), típicamente explicada como consecuencia del crecimiento demográfico, político o de la agricultura, pero insuficientes, «porque muchas sociedades a gran escala (p.ej. Egipto) combinan todos estos factores, pero no dieron lugar a religiones moralizantes». Es posible que la escalada en la prosperidad económica en esas tres regiones permitiera a la gente desprenderse de necesidades materiales como alimento, protección o afiliación, dado que estas podían ser mejor cubiertas por los emergentes sistemas de manejo de recursos. Estudios indican que «las personas subestiman el valor de una mayor riqueza o estatus cuando se cumplen estas necesidades, y dirigen su atención a otros dominios de las preferencias evolutivas, como la maximización del bienestar personal, el goce de la amistad, y el cultivo de la estética», todas metas típicas de los sistemas morales. Este parece ser el escenario que propicio la aparición de las religiones y los dioses moralizantes.
Nótese que estas nuevas preferencias involucran proyecciones y recompensas más elevadas, distantes o profundas, y abstractas, que de hecho encajan muy bien con el desarrollo de «la mente dopaminérgica» (Previc 2009). Según esto, el crecimiento social, el incremento del intercambio de bienes y logros materiales, la aparición de las naciones y el desarrollo de instituciones trascendentes, en conjunto alimentan la hiperagencia a su vez que necesariamente se soportan en niveles cada vez más abstractos de ésta, lo que crea un escenario propicio para la ideación de explícitos vigilantes sobrenaturales, y ulteriores sistemas de conceptos y toda suerte de construcciones lógicas sobre ellos (p.ej. doctrinas, teología). Entonces, retomando el asunto de que la expectativa de recompensa (regulada dopaminérgicamente) está presente tanto en la competencia intergrupal como en el castigo endogrupal, se debería hallar evidencia de otros efectos dopaminérgicos aquí asociados y vinculados a su vez a la hiperagencia, en cuanto a vigilancia sobrenatural se trata. Dando ya por descartado algún tipo de determinación neurobiológica para el sistema denominado religiosidad (R), se argumentará que, en contraste, la sensación de un vigilante invisible alrededor es otra manifestación del PM o hiperagencia, posible y probablemente controlada a un nivel neurobiológicamente mecánico.
4. Hacia una explicación neurobiológica de la vigilancia sobrenatural
Era necesario detenerse en este asunto del castigo sobrenatural porque, en contra de deducir que la R sea una adaptación por ello mismo, parece en cambio que estamos ante mecanismos cognitivos subyacentes que, si bien la R los canaliza muy efectivamente, no han aparecido por causa de ella: ya vimos que el Dios abrahámico, aunque paradigma de agente sobrenatural castigador (ciertamente un grupo exótico de entidades sobrenaturales-religiosas), es un desarrollo cultural bastante reciente. Bien, ahora veamos, de acuerdo con Johnson (2011 pp. 78-80), cuáles serían las razones por las que los agentes sobrenaturales castigadores son «los mejores disuasivos contra el egoísmo»:
- «las creencias sobrenaturales no son “cognitivamente costosas”»: pueden de hecho ser conductualmente costosas (p.ej. los rituales) pero la intuición mágico-sobrenatural es el «default cognitivo»;
- «Dios funciona porque Dios es “excesivo”»: de acuerdo a la ‘teoría de gestión de errores’, si el error de falso negativo (asumir que el fraude es secreto pero luego ser atrapado) es más costoso que el error de falso positivo (asumir que hay vigilancia y perderse los beneficios), entonces solo la hiperestimación de la probabilidad de detección (p.ej. creer que un agente sobrenatural vigila) va a evitar los peores errores; en tanto que «si los humanos están de hecho demasiado confiados acerca de sus probabilidades de irse haciendo trampa, como Schloss & Murray argumentan, entonces el sesgo de contrabalanceo tendría que ser aún mayor» (cf. Johnson 2009a);
- un sesgo cognitivo no religioso, asociado al exceso de confianza en evadir la detección, no puede ser simultáneo a un sesgo inverso (de baja confianza) —Dios es una alternativa;
- los sesgos cognitivos podrían actuar como advertencia, pero sufren de dos debilidades: (1) pueden suprimir el egoísmo, pero no las consecuencias, mientras que Dios reprime el egoísmo y castiga si el engaño es descubierto, (2) un sesgo cognitivo sólo afecta a las personas, sin validación externa, mientras que la creencia en el castigo sobrenatural es un aspecto compartido de la cultura que se ve reforzada por la comunidad entera y apoyada por las explicaciones de múltiples eventos;
- la evidencia empírica sugiere que la gente espera un castigo sobrenatural por un comportamiento egoísta (incluso los ateos en algunos casos), mientras que hay una falta de evidencia de mecanismos alternativos excluyentes de sobrenaturalidad (el solo exceso de confianza, como visto, trabaja en la dirección opuesta);
- cierta evidencia empírica sugiere que las creencias religiosas son «más eficaces» en la supresión de la conducta egoísta y promoviendo la cooperación, a diferencia de las creencias no religiosas equivalentes (p.ej. Sosis & Bressler 2003) —aún si los mecanismos alternativos existen en la actualidad o en el pasado, la religión puede haber tenido una ventaja competitiva;
- en tanto que Dios es un agente sobrenatural que trasciende las leyes naturales, ofrece una «mejor detección»: no es seguro que ocurra la detección del mundo real humano —de hecho, la extraordinariedad de los agentes sobrenaturales, como poder estar en varios lugares a la vez y tener acceso a la mente de las personas (cual ‘telépatas’), los convierte pues en vigilantes extraordinarios, en contraste con los vigilantes humanos;
- igualmente, sus facultades extraordinarias permiten a los agentes sobrenaturales castigar las consecuencias, que pueden escapar al castigo impuesto por humanos —aún la propia pena de muerte podría percibirse como un castigo finito, y en contraste p.ej. el castigo de Dios es infinito y puede de hecho ser más temido que la muerte: «ningún ser humano puede igualar la capacidad de castigo de Dios»;
- el castigo sobrenatural puede estar por sobre otras soluciones, aunque sean buenas, debido a una serie de limitaciones evolutivas (Johnson 2009b; McKay & Dennett 2009 —ver «LA EVOLUCIÓN DE LAS CREENCIAS FALSAS [I], [II], [III]»), como de tipo económica (el temor de la agencia sobrenatural podría haber sido biológicamente más barato o mejor para su selección o sostenimiento que las alternativas), o histórica (una capacidad para las creencias sobrenaturales podría haber estado más fácilmente disponible, especialmente teniendo en cuenta la evolución reciente de la ToM).5
- (1) primado conductual o acción ideomotora —las personas actúan generosamente en el contexto de atender a las palabras «Dios» o «profeta» precisamente porque tales están semánticamente asociadas a conductas prosociales; y,
- (2) vigilancia sobrenatural —tales palabras disparan la percepción de una vigilancia sobrenatural que incrementa la prosocialidad.
Empezaremos entonces por aclarar que lo así llamado ‘vigilancia sobrenatural’ no se reduce a una identidad con ‘Dios’ o ‘dios’, sino que remite a una agencia objetivamente carente de presencia física que es percibida como relativamente vigilante, o al menos que hace que el individuo se comporte como si se sintiera observado en sentido público (anecdóticamente: ¿acaso no sentimos como si ‘alguien’ nos pudiera estar observando precisamente cuando estamos realizando privadamente un engaño?). En este sentido, una vigilancia sobrenatural puede remitir a agentes imaginarios, abstractos y/o trascendentales pero no necesariamente religiosos, como p.ej. «la patria»; o bien puede remitir a un agente que sí es físicamente existente, pero que no es espacial/temporalmente inmediato al individuo, como «el Presidente». Para entender esto volvamos al concepto de «persona distribuida» de S. Guthrie: «el hecho que las personas, a través de sus artefactos distribuidos, típicamente son agentes en varios lugares al mismo tiempo. Ellas no necesitan ser visibles ni tangibles para tener una presencia». Así, la vigilancia sobrenatural por último remite a una circunstancia de percepción de lo que representa al agente o la mera intención de vigilar, como una imagen de un mirada o de ojos, que como se verá, puede alterar la conducta de engaño. Esto significa que la ToM funciona intrínsecamente como un ejercicio continuo de «sobrenaturalización» de los estímulos que recibe: un par de ojos retratados en una pared en realidad no están vigilando nada, sino que nuestra intuición social está así programada para reaccionar como si lo hicieran, e iniciar conductas al respecto, lo cual sería per se un asunto pues «sobrenatural»6 o «mágico». Entonces, debe haber evidencia de un mecanismo neurocognitivo, independiente del aprendizaje y del primado, que consista en modificar la conducta según la percepción de una vigilancia externa. Con esto podríamos responder la demanda de McKay & Dennett.
Se ha mostrado que no es un subproducto cultural la ‘atribución de intención al entorno’ o la sensación del ‘espacio (externo) como agente’ (la intrínseca sobrenaturalidad de la cognición social humana), de hecho señalada por Guthrie (2002) como un extendido ‘animismo’ en el reino animal más allá de la especie humana, lo que significa que «vigilancia» u «observación» son precisamente ‘contenidos mentales’ implícitos en tal atribución (de hecho como aspectos volitivos del agente). Tampoco se reducen a meras adquisiciones culturales, o por lo menos no completamente, las inferencias sociales, es decir la Teoría de la Mente (ToM), ni las proyecciones espaciotemporales de situaciones de interacción social, o sea el Mental Time Travel (MTT), disparadas por tal atribución. Entre tanto, todo esto tiene carácter ‘abstracto’: no trata sobre «personas», «situaciones» ni «reglas» específicas, sino que son patrones perceptuales (unos pocos estímulos sensoriales disparan un limitado juego de inferencias) y categoriales (tales inferencias responden a «conceptualizaciones» automáticas: si un objeto inanimado tiene cierta motricidad o apariencia entonces es categorizado como animado) de tipo generalista, funcionales tanto p.ej. para cualquier persona (si real o imaginada —p.ej. como una «presencia»), objeto o evento (que sabemos igualmente se les puede atribuir animación y así desencadenar las inferencias ToM/MTT).
Por supuesto, es un hecho la influencia de factores culturales, y extra-genéticos (p.ej. embarazo, alimentación, enfermedades), como tampoco la ToM ni el MTT están ya presentes al nacimiento: se les está caracterizando como (neuro)biológicos por el hecho de que el diseño y la construcción de su neuroarquitectura y regulación neurofisiológica, aunque no presentes al nacimiento, responden a la activación de un número de genes regulando un plan de migración neuronal, formación de conexiones a pequeña y gran escala (sinapsis, materia blanca: ver «Teoría de la Mente y neurodesarrollo»), y el perfil de neurotransmisores/hormonas, todo lo que co-determina la capacidad de percibir, categorizar e inferir. Aunque ciertamente el desarrollo óptimo se da en la medida en que el cerebro interactúa con los estímulos y el entorno, está claro que la neurogenética juega un papel de peso: consistentemente, los cerebros autistas más o menos expuestos a los mismos estímulos externos tienen sin embargo impedimentos en cognición social, precisamente por anomalías neurales (p.ej. Dapretto et al. 2006; Sasamoto et al. 2011: «reducción de materia gris en el área cortical rodeando el surco temporal superior izquierdo —STS—, el cual es considerado de importancia en la percepción social»). Estas capacidades, centralmente la hiperagencia (‘atribución de mentalidad al entorno’, intuición teleológica, PM), predisponen fuertemente a la elaboración de narrativas y creencias sobrenaturales, pero nada aquí implica que estas capacidades en sí mismas contengan preprogramado un concepto tal como «Dios castiga». De hecho las anomalías agentivo-sociales de los autistas (Síndrome de Asperger) los hace ciertamente ajenos a Dios (Scientific American, 2010; cf. Heywood & Bering 2010; Caldwell-Harris et al. 2011) —la misma agencia, «el entendimiento de las metas y los planes de otros» es deficiente en ellos (Gray et al. 2011 p. 478 § 5).

Haley & Fessler (2005, ver aquí o aquí) mostraron que la sola presencia de un gráfico estilizado de dos ojos (p. 250 Fig. 1, ver arriba), correlaciona con la duplicación de la generosidad, aún en anonimia. Los autores: «si, como creemos, los sistemas afectivos motivando el comportamiento prosocial sirven al objetivo final de crear y mantener relaciones sociales individualmente beneficiosas [...] entonces la percepción y la toma de decisión humanas deben ser especialmente sensibles a las señales de la presencia de otros. En particular, los seres humanos están innatamente a tono con la presencia de caras, y, como estímulos, los ojos juegan un papel importante en este sentido [...], probablemente porque los ojos frente a uno han servido como indicadores fiables de la atención dirigida hacia uno mismo, en el transcurso de la evolución humana y en los típicos contextos modernos» (ibid. p. 254 § 2). Que el reconocimiento facial sea un mecanismo automático e innato, neuro-categorialmente «especificado en (no) más detalles que tres áreas de alto contraste» (Batki et al. 2000), es algo que se considerará como un hecho (ver «RECONOCIMIENTO FACIAL: UNA HABILIDAD ESPECÍFICA Y HEREDABLE»), y consistentemente, la atención hacia los ojos debe ser un asunto implícito en ello (Itier & Batty 2009), de carácter igualmente adaptativo: «reconocer que eres el foco de atención de otro organismo sirve como un sistema de alerta temprana de que el otro animal puede tener como objetivo atacarte, o estar interesado en ti por alguna otra razón» (Batki el al. 2000 p. 4 § 3). Entre tanto, nótese que esta ilustración nos remite a la interacción entre los módulos de reconocimiento facial, agencia y ToM, cuya propia interacción, conformada por la materia blanca y su configuración neuroanatómica, es también un proceso más genética que ambientalmente influido. Por supuesto, Batki et al. (2000) reportaron el significativo mayor tiempo que los neonatos (un centenar de ellos, con un promedio de 36 horas de nacidos) prestan atención a los ojos abiertos, en contraste con a los cerrados.
- (1) su temprana manifestación —además del reporte de los autores evidenciando lo extremadamente temprano del mecanismo, a los 2 meses los bebes muestran preferencia respecto a los ojos sobre otras regiones del rostro, de 3 meses dirigen su mirada hacia un objetivo que es mirado por un adulto, a los 4 meses discriminan el evitamiento de la mirada, y de 5 meses prefieren los rostros que muestran contacto visual directo;
- (2) el automatismo de la inspección ocular en adultos —que a pesar de estar advertidos de no ver ciertas señales, no pueden evitar dirigir la mirada hacia ellas;
- (3) la evidencia neurofisiológica animal —se ha identificado una población neuronal en el surco temporal superior (STS) «que responde al máximo a la dirección particular que los ojos de otro mono buscan o su cabeza apunta», como también respuestas eléctricas en la piel y activación medular en monos;
- (4) la evidencia de neuroimagen en humanos —confirma el rol del STS en la percepción de la mirada; y,
- (5) los estudios de lesión humanos/monos —daños en el STS de ambos primates producen deficiencias en el reconocimiento de la mirada (ver también Emery 2000).

Entonces, lo que aquí tenemos es una efectiva relación neuroanatómica y funcional del «Núcleo de Agencia», el fundamento de la cognición social-afectiva STS-amígdala-CFO, con la red CFO-frontomedial-amígdala-CCA de la toma de decisión, que incluye al CPFDL. Hay que recordar que el córtex prefrontal medial (CPFM) conforma parte constitutiva de la agencia y de la ToM (ver Fig. 1 ver abajo; ver «Agencia abstracta sin ToM, y alcance de la ToM hasta la agencia sobrenatural–religiosa»), junto con el STS y su región posterior inmediata, la junción temporoparietal (JTP). Ambas regiones CPFM/JTP están implicadas en la ‘mentalización’ o atribución de agencia, pero no tienen la misma función: para Saxe et al. (ver la referencia sobre agencia abstracta sin ToM) la JTP sostiene la específica atribución de mentalidad, de hecho identificando/categorizando automáticamente algo como ‘un agente’ vía STS-MB (reconocimiento de movimiento biológico e imputación de intención a partir de ello), mientras el CPFM representa la información social/emocional del agente. De hecho, a diferencia del sistema STS-JTP, dedicado a la identificación/categorización peceptual, el CPFM conforma un sistema de integración cognición/emoción, evaluación y toma de decisión, en virtud de sus directas conexiones con el CPFDL (modulación emocional, repuesta motora, narrativa), el CCA (detección de errores), la amígdala (reacción emocional), y la sustancia negra (ganglios basales: recompensa/dopaminergia). Frith (2007) discute si es que el CPFM es realmente necesario para mentalizar, ya que p.ej. a pesar de la evidencia a favor, hay un caso «excepcionalmente raro» de daño específico de tal región sin perjuicio de ejecutar la mentalización (Bird et al. 2004). Es posible, como los autores discuten, dado que el daño no involucró absolutamente todo el CPFM, que aún haya subregiones funcionando óptimamente, o bien pudo haberse dado algún tipo de recuperación anatómica, o por último, dado que la mentalización o ToM implica las ya conocidas regiones JTP, STS, OFC, el daño del CPFM no afecte sustancialmente su ejecución.
A esto hay que añadir lo señalado por nuestra parte sobre que, según lo observado sobre el neurodesarrollo infantil, la mentalización puede de hecho ser previa a la maduración del CPFM y de la región frontopolar (aún más tardía en madurar, ya en un lapso de 3 décadas), tenida esta última como sustrato de los altos procesos de pensamiento abstracto, en tanto que atribuir mentalidad o categorizar algo como un agente supone una casi extrema abstracción cognitiva. Nótese por tanto que el STS categoriza agencia a partir tan solo del movimiento biológico y/o del reconocimiento de ojos/rostro. Los neonatos humanos reconocen ojos, rostros y forma humana (ver «ANIMISMO, FOLK BIOLOGY Y LA PREDISPOSICIÓN AL MOVIMIENTO BIOLÓGICO»), mientras los infantes de 6.5 meses atribuyen agencia a un objeto inanimado (Csibra 2008) y de 1 año de edad categorizan como ‘agente’ a algo que tenga ojos o responda interactivamente (Johnson et al. 1998), todo esto parece que no necesariamente debido a la maduración de ciertas regiones prefrontales medial/polar, en tanto que están empezando a desarrollarse a esa edad, sino por hiperactividad STS-inferotemporal durante esta etapa del desarrollo. De hecho, se ha mostrado actividad en la región temporal superior posterior y parietal inferior (o sea abarcando la JTP), en bebés nacidos prematuramente a los 7 y 8 meses de gestación, en un estudio de las capacidades innatas para el lenguaje (Mahmoudzadeh et al. 2013). Todo esto podría facilitar que los infantes a partir de los 3 años sean capaces de inventar amigos imaginarios (Gleason & Hohmann 2006). Considerando a Saxe y a Frith por un lado, y observando el neurodesarrollo infantil por otro, la abstracción agentiva resulta en una propiedad intrínseca y muy temprana del STS.
Con todo esto lo que se pretende es una tentativa explicación neurobiológica de la sensación de sentirse observado, y ciertos experimentos podrían ilustrarla. En un grupo de niños entre 5 y 9 años de edad, los sujetos modularon su conducta de engaño (hacer trampa) cuando creyeron en la presencia de un agente invisible que les observaba, desconocido e inventado (la «Princesa Alicia»), para una situación de juego cuyas reglas serían casualmente observadas por ella (Piazza et al. 2011). Lo que tenían que hacer los niños era lanzar bolas a un blanco, con los aciertos siendo premiados; mientras que se elaboraron tres situaciones: una de control, sin vigilancia de ningún tipo (con desconocimiento de Alicia), con la presencia de un adulto, y con la presencia del agente invisible (ver infra). Las ganas de hacer trampa fueron inhibidas casi igual con la presencia de un adulto o de Alicia, y los menores fueron los más creyentes en ésta y los más inhibidos de engañar, mientras que los niños «escépticos» (típicamente mayores) de esta presencia, aunque hicieron trampa, verificaron que la entidad no estuviera presente. Para los autores esto «demuestra que los efectos prosociales de los pensamientos primados sobre agentes sobrenaturales no están limitados a los conceptos religiosos culturalmente fundados (p.ej. Dios, espíritus ancestrales) sino más bien están generalizados hacia agentes sobrenaturales novedosos a los cuales los niños no han sido expuestos previamente» (ibid. p. 8). Interesantemente, este no es un asunto de cognición infantil, sino que existe «evidencia experimental acumulada de que las creencias sobrenaturales promueven el altruismo y el seguimiento de las reglas en los adultos» (ibid. p. 2).

«Arreglo experimental (como visto con una cámara oculta en un altavoz montado). El objetivo para las bolas está en la pared del fondo. La línea blanca en el suelo era la que los participantes recibieron instrucciones de no cruzar al lanzar. La silla en el extremo derecho es de la Princesa Alicia. En esta escena, un niño inspecciona la silla para obtener evidencia de la presencia de la Princesa Alicia.» (Imagen de Piazza et al. 2011 p. 4).
5. Discusión
En cuanto al asunto genético se puede descartar una codificación específica para la religión, mientras que los siguientes aspectos tienen indicios de base genética o bien ya cuentan con genes identificados:
- el pensamiento mágico → genes dopaminérgicos;
- la prosocialidad y los aspectos socio-afectivos → genes oxitoninérgicos, vasopresinérgicos, serotoninérgicos;
- la neuroanatomía correspondiente → genes del área fusiforme de las caras, de la junción temporoparietal, de la RpD.
5.1. ALGORITMOS COMPUTACIONALES Y JERARQUÍA DINÁMICA NEURO-CULTURAL
- los algoritmos computacionales de la percepción conceptual, que son automáticos, mínimos y específicos, como el reconocimiento de movimiento biológico, de rostros, de ojos.
- la agencia (y así a la teleología, la causalidad, el mentalismo, el viaje mental en el tiempo, al animismo),
- los procesos finales de interacción de las emociones, la memoria, la toma de decisión, y el pensamiento analítico.
Se estima que la sugerencia de un sustrato biológico para el PM es robusta, aunque aparentemente no existe literatura específica al respecto. El acercamiento desde la biología ha sido indirecto: tenemos interesantes progresos en neurociencia y ciencia cognitiva sobre las bases neurales asociadas a aspectos religiosos como la oración, hablar en lenguas, las creencias comunes sobre Dios, la autotrascendecia y la experiencia mística, con lo que prácticamente se cubre todo el espectro ideacional de la religión; adicionalmente se han estudiado los sustratos neurales de ciertos estados alterados no necesariamente religiosos como la experiencia extracorporal, las sensaciones de presencia extraña y las alucinaciones. Desde la psiquiatría tenemos la investigación sobre la esquizotipia, la esquizofrenia, y el autismo. Todos en suma, fenómenos religiosos y no religiosos, normales o patológicos, son bien conocidos por involucrar la agencia y el PM, por lo que cualquier hallazgo neurológico y fisiológico en esos campos enriquece la explicación de éstos. En principio, o al menos en una fase inicial del proceso de computación neural, los sustratos de la agencia serían los mismos para el PM, y lo que se conoce hasta la fecha es que ese inicio está en un conjunto de mecanismos que comprende el reconocimiento de movimiento biológico, el reconocimiento facial, el reconocimiento de mirada, y el reconocimiento de intención. A estos los denominamos núcleo de agencia. El proceso se extiende hacia la red de la Teoría de la Mente, el Mental Time Travel, la Toma de Decisiones, y en una última fase el filtro analítico. Esto se denominará jerarquía dinámica neuro-cultural, y puede explicar no sólo la diferencia entre el pensamiento mágico (mayor influencia biológica) y la creencia mágica (mayor influencia cultural), sino la posibilidad de inhibición analítica de la cascada inferencial agentivo-mágica.

Fig. 3. Esquema de los algoritmos computacionales y sus correspondencias neurales aproximadas, más un diagrama simple de la jerarquía dinámica neuro-cultural. Este gráfico resume los resultados de los tres ensayos enfocados en la neurobiología del pensamiento mágico. La evolución del cerebro social definió los algoritmos de la percepción: todo puede ser agencia intencional. El principio básico es poseer una propia realidad interna a partir de conceptos socialmente relevantes, pero que pueden no coincidir con la realidad externa siempre: proyectar intenciones y patrones a la realidad externa, asumir entidades intencionales como causas y construir escenarios imaginarios contrafactuales. Así, es una cognición mágica y sobrenaturalista que forma la sensación de cohesión intuitiva y significativamente emocional entre individuos: agentividad mágica. (Hacer click en la imagen para ampliarla).

Fig. 4. Diagrama de la jerarquía dinámica neuro-cultural con dos ejemplos: los procesos implícitos/inconscientes de la vigilancia sobrenatural y el reconocimiento facial, resultando en creencias explícitas/conscientes culturalmente perfiladas. Las líneas gruesas indican conexión robusta, y las líneas punteadas conexión débil o relativa. Esto refiere a las situaciones en las que p.ej. bajo condiciones de respuesta veloz/inadvertida, o patológicas, la agentividad y el pensamiento mágico dominan el lenguaje (aún si conscientemente nos declaramos descreyentes o escépticos). (Hacer click en la imagen para ampliarla).
Finalmente, y bajo el enfoque holístico de que el PM, la agencia, el teleologismo, y el animismo son conceptos diferentes que refieren a un mismo núcleo cognitivo, hay en la literatura un interesante concepto adaptacionista del PM: la «economía cósmica de compartir» en el animismo de las sociedades cazadoras-recolectoras. La idea es como sigue: dado que en este animismo omnipresente todo está conectado con todo, y todo «comunica» significados de naturaleza social hacia el hombre, entonces todo lo que el entorno «ofrece» a las personas debe ser compartido y dado a los congéneres físicos o espirituales (Bird-David 1992, resumen / completo). Este análisis antropológico del animismo es consistente con la psicología ultrasocial del PM, pero sobre todo, tal cosmovisión altruista podría explicarse por la neurobiología de la prosocialidad a partir de la detección de agentes y la básica percepción de ‘el espacio (externo) como agente’. Quizás el PM pueda ser o haya sido un estilo cognitivo adaptativo de la evolución humana. Las interacciones encontradas entre oxitocina (OT) y dopamina (DA) involucradas juntamente en las inferencias mentalísticas y el apego hacia recompensas abstractas, además obviamente de la propia dimensión de apego social en actividades colectivas como la caza y la recolección, tentativamente, parece una clara explicación neurofisiológica del PM como promotor y/o producto del apego social y lo que ello implique como el altruismo y la generosidad. Si tal como Guthrie desde la antropología o Subbotsky desde la psicología nos descubren el animismo y el PM, estos son pues (sub)productos de la evolución de nuestra ultrasociabilidad y por tanto a la vez promotores de ella.
La biología de la agentividad mágica luce clara: los humanos estamos predispuestos a ella, muy a pesar de quedar fuera de contexto en las sociedades técnico-científicas, porque, simplemente, es el principio cohesivo entre humanos. El reconocimiento biológico, facial, de mirada, y de intención, conforman el núcleo de agencia, que en traslape con el viaje mental en el tiempo (MTT), son un sistema funcional permanente de proyección imaginaria de agentes y escenarios. Es un principio computacional que permite, por ejemplo, que el lenguaje tenga sentido: las palabras y las ideas son representaciones de objetos tangibles pero también de cosas puramente abstractas, que sin embargo se perciben como reales, y es necesario que así sea para que, en otros ejemplos, la matemática, la justicia y la libertad funcionen. Vemos una equivalencia conceptual entre esto y lo que precisamente define al pensamiento mágico: conexiones, causas e intenciones que objetivamente pueden no existir en el entorno, y de ahí que se hable de la agentividad mágica. Queda claro que sin esto no se hubiera forjado la cultura ni la filosofía, y paradójicamente, por debajo de sus diferencias, ni la ciencia misma. En último término, la agentividad mágica caracteriza la inteligencia humana en tanto somos un primate social.
1. Siempre apelando a la definición dada en el marco de la investigación científica cognitiva: cf. «EXPLICAR LA RELIGIÓN: UNA CLAVE PARA ENTENDER LO HUMANO. UNA DEFINICIÓN CIENTÍFICAMENTE OPERATIVA». P.ej. ambas definiciones de Atran & Norenzayan (2004) y Boyer & Bergstrom (2008) podrían reducirse a dos aspectos generales presentes «en todas las sociedades» (mejor dicho, religiones): (1) «demostraciones comunales apasionadas de compromisos costosos», que las personas manifiestan como conductas y creencias respecto a los (2) «mundos contraintuitivos gobernados por agentes sobrenaturales». Para efectos de identificar dos mayores aspectos de la religión aquí se asume que, (1) podría caracterizarse como R, y (2) como ‘creencia mágica’ (que ya vimos antes no es exactamente lo mismo que PM). Ciertamente, habiendo visto en la PARTE 2 cómo el PM impregna o subyace a los más diversos aspectos de la interacción social, la organización institucional, el afrontamiento de el estrés y la percepción del mundo natural, sería más propio asumir que el PM, como ‘estilo cognitivo por defecto’, está involucrado estructuralmente en ambos aspectos. De hecho la ‘creencia mágica’ (p.ej. supersticiones y creencias paranormales, místico-religiosas y pseudocientíficas) se trata de una caracterización del PM, que en sí misma es fundamental para la R.
2. «La palabra prosocial no aparece en la mayoría de los diccionarios, fue creada por los científicos sociales como un antónimo de antisocial. La conducta prosocial abarca un amplio abanico de acciones destinadas a beneficiar a una o más personas distintas de uno mismo, conductas como la de ayudar, consolar, compartir y cooperar. La palabra altruismo ha sido a veces utilizada para referirse a un subconjunto de estos comportamientos, por ejemplo, la ayuda autosacrificada o ayudar en ausencia de obvias recompensas externas. Tal uso parece inadecuado, entre tanto, porque el altruismo es un concepto sobre motivación. El altruismo es la motivación para aumentar el bienestar de otra persona; es contrastado con el egoísmo, la motivación para aumentar el propio bienestar (MacIntyre, 1967). No hay una correspondencia uno-a-uno entre la conducta prosocial y el altruismo. La conducta prosocial no necesita ser motivada por el altruismo; la motivación altruista no tiene por qué producir una conducta prosocial.» (Batson & Powell en Millon, Lerner & Weiner 2003 p. 463 § 1).
3. El rol del córtex temporal ha sido de hecho tratado más extensivamente en «NEUROCIENCIA DE LA RELIGIÓN (V): CUANDO LAS INTENCIONES DEL ENTORNO TIENEN ROSTRO», y «NEUROCIENCIA DE LA RELIGIÓN (VI): EL PAPEL DEL LÓBULO TEMPORAL».
4. En el campo de la cognición social, un polizonte es «quien disfruta de los beneficios del grupo creado por las contribuciones de otros sin pagar los costos» (Henrich 2006).
5. Nótese que, si realmente existe tal sesgo cognitivo de agencia y castigo sobrenatural, que sea ciertamente un default cognitivo, esto podría dar cuenta de la posibilidad que un teísta aún llegando a reconocer que Dios, el Yahvé del Antiguo Testamento, sea un castigador implacable, sea a su vez necesario.
6. Tal equiparación está justificada semánticamente (para ello las referencias a la RAE), y es algo previamente argumentado y discutido: ver aquí, y aquí.




No hay comentarios:
Publicar un comentario